
Энергия в экосистемах и продуктивность экосистем
Энергия в экосистемах и продуктивность экосистем
1.1.1. Поток энергии в экосистемах и жизнь как термодинамический процесс
Жизнь есть особая форма существования и движения материи, высшая по отношению к физической и химической формам. Все разнообразие проявлений жизни сопровождается превращениями энергии, хотя энергия при этом не создается и не уничтожается. С точки зрения процессов преобразования энергии природная экосистема может рассматриваться как любая физическая система, для которой должен выполняться первый закон термодинамики, или закон сохранения энергии, гласящий, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново. Энергия, получаемая в виде света поверхностью Земли, уравновешивается энергией, излучаемой с поверхности Земли в форме невидимого теплового излучения.
Сущность жизни состоит в непрерывной последовательности таких изменений, как рост, самовоспроизведение и синтез сложных химических соединений. Без переноса энергии, сопровождающего все эти изменения, не было бы ни жизни, ни экологических систем.
Непрерывный поток солнечной энергии, воспринимаясь молекулами живых клеток, преобразуется в энергию химических связей. Создаваемые таким образом (например, при фотосинтезе) химические вещества последовательно переходят от одних организмов к другим: от растений к растительноядным животным, от них – к плотоядным животным первого порядка, затем второго порядка и т.д. Этот переход рассматривается как последовательный упорядоченный поток веществ и энергии.
Если температура того или иного живого тела выше температуры окружающей среды, тело будет отдавать тепло до тех пор, пока его температура не сравняется с температурой окружающей среды. В конечном итоге энергия любого живого тела может быть рассеяна в тепловой форме, после чего наступает состояние термодинамического равновесия, и дальнейшие энергетические процессы оказываются невозможными. О такой системе "тело-среда" говорят, что она находится в состоянии максимальной энтропии.
Энтропия – мера количества связанной энергии, которая становится недоступной для использования. Этот термин также используется как мера изменения упорядоченности. Понятие энтропии служит основой для формулировки второго закона термодинамики или закона энтропии: процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что энергия переходит из концентрированной формы в рассеянную (деградирует), т.е. энергетические процессы идут в сторону возрастания энтропии.
Отличительной особенностью живых организмов является способность выполнять работу против уравновешивания с окружающей средой за счет образования сложно организованных упорядоченных молекулярных структур. Физики давно были обеспокоены тем фактом, что сохранение функциональной упорядоченности живых существ как бы опровергают второй закон термодинамики. Илья Пригожин, получивший Нобелевскую премию за работы по неравновесной термодинамике (1962 год), разрешил это кажущееся противоречие, показав, что способность к самоорганизации и созданию новых структур может встречаться в системах, далеких от равновесия и обладающих хорошо развитыми диссипативными структурами, откачивающими неупорядоченность. Дыхание высокоупорядоченной биомассы можно рассматривать как диссипативную структуру экосистемы.
Таким образом, важнейшая термодинамическая характеристика организмов, экосистем и биосферы в целом – способность создавать и поддерживать высокую степень внутренней упорядоченности, т.е. состояние с низкой энтропией. Процессы в экосистемах идут по пути постоянного и эффективного рассеяния легко используемой энергии (например, энергии света или пищи) и превращения ее в энергию, используемую с трудом (например, в тепловую). Упорядоченность экосистемы, т.е. сложная структура биомассы, поддерживается за счет дыхания всего сообщества, которое постоянно "откачивает из сообщества неупорядоченность". Таким образом, экосистемы и организмы представляют собой открытые неравновесные термодинамические системы, постоянно обменивающиеся с окружающей средой энергией и веществом, уменьшая этим энтропию внутри себя, но увеличивая энтропию вовне в согласии с законами термодинамики.
Рекомендуемые материалы
Экология, по сути дела, изучает связь между светом и экосистемами и способы превращения энергии внутри системы, поток энергии в экосистеме. Особое внимание экологов привлекают вопросы преобразования энергии горючего, атомной энергии и других форм концентрированной энергии в индустриализованном обществе.
Для описания "поведения" энергии в экосистемах подходит понятие "поток энергии", поскольку в отличие от циклического движения веществ превращения энергии идут в одном направлении. Источником энергии служит Солнце. Попав на Землю, лучистая энергия Солнца стремится превратиться в тепловую. Лишь очень небольшая часть световой энергии, поглощенной зелеными растениями, превращается в потенциальную энергию пищи, большая же ее часть (99 %) превращается в тепло, покидающее затем и растение, и экосистему, и биосферу. Весь остальной живой мир получает необходимую потенциальную химическую энергию из органических веществ, созданных фотосинтезирующими растениями или хемосинтезирующими микроорганизмами. Животные поглощают химическую потенциальную энергию пищи и большую ее часть переводят в тепло, а меньшую вновь переводят в химическую потенциальную энергию заново синтезируемой протоплазмы. На каждом этапе передачи энергии от одного организма к другому значительная ее часть рассеивается в виде тепла.
1.1.2. Универсальная модель потока энергии
Данная модель может быть приложена к любому живому компоненту системы, будь то растение, животное, микроорганизм, особь, популяция или трофическая группа.
Общее поступление энергии будем обозначать буквой I. Для облигатных (100%) автотрофов – это свет, для облигатных гетеротрофов – органическая пища. Возможно и использование обоих источников энергии. Не вся энергия, поступающая в биомассу В, подвергается превращению. Потери обозначены как NU. Таким образом, ассимилируемая часть энергии А равна I-NU. Эффективность ассимиляции А/I =1 - NU/I принимает значения в широких пределах от 0 до 1. Оно может быть очень мало, как в случае усвоения света растениями или ассимиляции пищи у животных – детритофагов, или же очень велико, как в случае животных или бактерий, питающихся высококалорийной пищей, например сахарами и аминокислотами.
Для автотрофов ассимилированную энергию принято называть валовой первичной продукцией или валовым фотосинтезом. Для гетеротрофов А – это пища или "метаболизированная энергия".
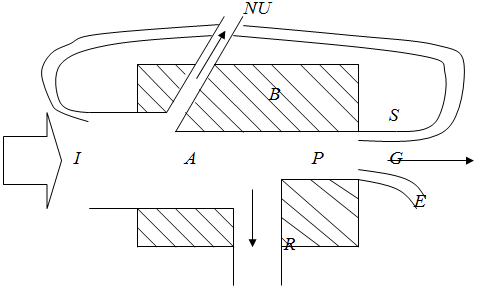
I – поступившая или поглощённая энергия;
NU – неиспользованная энергия;
A – ассимилированная энергия;
P – продукция;
R – дыхание;
B – биомасса;
G – рост;
S – накопленная энергия;
E – выделенная энергия;
A = I – NU ; P = A – R ; G = P – S – E.
Ассимилированная энергия А делится на продукцию системы Р и на дыхание системы R. Та часть ассимилированной энергии А, которая окисляется и теряется в форме тепла, называется дыханием (R), а та часть, которая превращается в новое или принадлежащее другому виду органическое вещество, называется продукцией (Р). У растений это чистая продукция, у животных – вторичная продукция .
Продукция может принимать различные формы: G – рост или увеличение биомассы, Е – ассимилированное органическое вещество, выделяемое с экскрементами, секретируемое или извлекаемое, S – запас, например, жировые накопления, которые могут быть использованы позже. Обратный поток S можно назвать и "рабочей петлей", поскольку это та часть продукции, которая используется для обеспечения энергией в будущем, например, хищник использует энергию запасенных веществ, чтобы найти новую жертву.
Продукция экосистемы Р увеличивает биомассу экосистемы В. Чтобы выявить связь между потоком энергии и биомассой, последнюю принято представлять в единицах энергии: в калориях или джоулях. И это вполне естественно. Любое количество органического вещества эквивалентно некоторому количеству энергии. Эту энергию можно извлечь, разрушив химические связи органического вещества. 1г сухого органического вещества растения в среднем соответствует 18.7 кДж энергии (4.5 ккал). В более богатых белками и жирами семенах растений энергетический эквивалент выше – около 20.0 кДж (5.2 ккал). 1г сухого вещества позвоночного животного (мяса) заключает 23.5 кДж (5.6 ккал).
Важнейшей энергетической характеристикой экосистемы является отношение Р/R или R/B. В экосистеме отношение общего дыхания сообщества к его суммарной биомассе (R/B) можно рассматривать как отношение затрат энергии на поддержание жизнедеятельности к энергии, заключенной в структуре, или как меру термодинамической упорядоченности. Данную величину называют "соотношением Шредингера” и оно служит мерой экологического оборота. Это отношение (R/B) равно отношению прироста энтропии (и соответствующей работы), связанного с поддержанием структуры, к энтропии упорядоченной части. Чем больше биомасса, тем больше затраты на поддержание; но если размер единиц, на которые поделена биомасса, достаточно велик, то затраты на поддержание процессов, идущих против температурного градиента, в пересчете на структурную единицу биомассы будут ниже. Величина Р, напротив, сравнительно велика в активных популяциях мелких организмов, например бактерий или водорослей, в молодых, быстро растущих сообществах и в системах, получающих энергетические дотации. При стрессовых воздействиях на систему R возрастает.
1.1.3. Энергетические характеристики биосферы
На биосферу из космоса падает солнечный свет с энергией 2 кал/см2 мин (так называемая «солнечная постоянная »). Но, проходя через атмосферу, он экспоненциально ослабляется; в ясный полдень до поверхности Земли может дойти не более 67% энергии Солнца, то есть 1.34 кал/см2 мин. Ещё более ослабляет интенсивность солнечного света облачный покров, вода и растительность. В итоге, поступление солнечной энергии к автотрофному слою экосистемы за день варьирует от 100 до 800 кал/см2 (в среднем 300-400 кал/см2). Неоднородно поглощение биосферой солнечного света и по частоте. Растительность сильно поглощает синие и красные лучи, а также дальнее инфракрасное излучение; зелёный свет поглощается не так сильно, ближнее инфракрасное излучение – очень слабо.
Попадаемая в биосферу солнечная энергия делится в следующей пропорции: 30% - отражается, 46% - прямо превращается в тепло, 23% - обуславливают испарение и осадки, 0.2% - тратится на ветер, волны, течения, 0.8% - идёт на фотосинтез.
Итак, большая часть биосферы получает ежедневно около 3000 ÷ 4000 ккал/м2 или 1.1÷1.5 млн. ккал/м2 в год. Из них в валовую первичную продукцию автотрофов переходит максимум 50 тыс. ккал/м2 в год, в среднем для биосферы – 2 тыс. ккал/м2 в год; в чистую первичную продукцию переходит максимум 40 тыс. ккал/м2 в год, в среднем для биосферы – 1 тыс. ккал/м2 в год.
1.1.4. Концепция энергетической субсидии
Высокая продуктивность и высокое отношение чистого урожая к валовому поддерживаются ценой больших вложений энергии, затрачиваемой на обработку земли, орошение, удобрение, селекцию, борьбу с вредными насекомыми. В горючем, которое расходуется сельскохозяйственными машинами, заключено не меньше энергии, чем в солнечных лучах, падающих на поля. В США вклад энергии топлива в сельское хозяйство увеличился с 1900 по 1970-е годы в 10 раз, примерно с 1 до 10 калорий на каждую калорию полученной пищи.
Сельское и лесное хозяйство, животноводство и т.п. требуют огромных потоков дополнительной энергии, которая выполняет немалую часть работы, в естественных условиях производящейся за счёт самой системы. Естественно, что при появлении этой дополнительной поддержки виды, входящие в естественную систему, оказываются неприспособленными к новой ситуации; поскольку их генетическая программа заставляет их по-прежнему выполнять всю работу, никакого выигрыша не получается. Но виды, не приспособленные к «самообслуживанию», в таких условиях получают преимущества, и им благоприятствует как искусственный, так и естественный отбор. Далеко зашедшее одомашнивание превращает организмы в «живые машины для производства органики ». Работа этих организмов по самоподдержанию заменяется работой сельскохозяйственных механизмов и машин, направляемых человеком. Таким образом, большая часть энергии при производстве современной сельскохозяйственной продукции берётся не от Солнца, а из ископаемого топлива.
Всякий источник энергии, уменьшающий затраты на самоподдержание экосистемы и увеличивающий ту долю энергии, которая может перейти в продукцию, называется вспомогательным потоком энергии, или энергетической субсидией.
Виды, приспособленные к наличию энергетической субсидии, резко снижают свою производительность в отсутствии последней. Поэтому развивающиеся страны не смогли воспользоваться достижениями «зелёной революции », произведёнными в развитых странах.
“Зелёная революция” была вызвана выведением путём селекции новых сортов сельскохозяйственных культур с высоким отношением пищи к волокну, приспособленных к тому, чтобы хорошо реагировать на массивные субсидии в форме энергии, орошения и удобрения. Без этих поступлений “чудесный рис” и другие новые сорта дают урожай ниже, чем традиционные сорта, не требующие таких субсидий.
Важен также и уровень субсидии. Малая субсидия также плоха, как и большая. Существует оптимальный уровень субсидии, отклонения от которого приводит к снижению производительности.
Источник энергетической субсидии может быть как естественного характера, так и антропогенного характера. В естественных условиях естественные сообщества, получающие добавочную энергию от природных её источников, дают наибольшую валовую продуктивность. Так, например, участок побережья, получающий в виде приливных или других потоков воды оптимальную энергетическую субсидию, имеет примерно такую же валовую продуктивность, как и интенсивно возделываемое кукурузное поле. Указанная энергетическая субсидия замещает часть энергии, затраченной на дыхание и другие функции, поддерживающие целостность системы, в результате чего эта энергия идёт на перенос минеральных веществ и перемещение пищи и отходов.
Как правило, по валовой продуктивности культурные экосистемы не превосходят некоторые природные. Конечно, человек увеличивает валовую продуктивность, доставляя воду и питательные вещества туда, где они служат лимитирующим фактором (в пустынях). Но больше всего человек увеличивает чистую продуктивность сообщества, направляя туда дополнительную энергию и уменьшая тем самым расход продукции на автотрофном и гетеротрофном уровнях (и увеличивая тем самым урожай на потребу себе самому).
1.1.5. Использование первичной продукции человеком
Удельная по площади валовая продуктивность меняется от экосистемы к экосистеме в диапазоне от 200 до 20000 ккал/м2год, а общая валовая продукция Земли имеет порядок 1018 ккал/год.
Каждому человеку для обеспечения его жизнедеятельности требуется около 1 млн. ккал, т.е. для поддержания так называемой «человеческой биомассы» в 6 млрд. человек требуется 6´1015 ккал энергии пищи. А из указанной валовой продукции Земли удаётся ежегодно собирать 6.7´1015 ккал пищи, но из-за плохо налаженного распределения, потерь и низкого качества части урожая этого оказывается недостаточно.
Примерно 30% всего человечества живёт в развитых странах, валовой национальный продукт (ВНП) на душу населения которых более 1000 долл.. Около 65% человечества живёт в развивающихся странах, где ВНП на душу населения меньше 300 долларов, а обычно меньше чем 100 долларов. Скорость прироста населения в развитых странах низок (менее 1%), а в развивающихся – велик (более 2%). Для развивающихся стран характерна низкая продукция в пересчёте на гектар, так как отсутствие средств не позволяет им вкладывать в сельскохозяйственные энергетические субсидии, необходимые для получения высоких урожаев. Таким образом, человечество разделено резко на две части, лишь 5% живёт в «среднеразвитых» странах.
Из-за преобладания населения развивающихся стран средний мировой сбор зерновых культур гораздо ближе к нижнему, чем к возможному верхнему пределу. В развивающихся странах урожаи растут медленнее, чем численность населения; они вынуждены ввозить продукты питания во всё больших объёмах. Например, для хлебных злаков разрыв в продуктивности в 1969 году составлял 16.3 центнеров/га, а в 1977 году вырос до 17.4 центнеров/га. За тот же период пахотные площади во всём мире выросли примерно на 15%, но в Европе, США и Японии они сократились. Развивающиеся страны вынуждены увеличивать площади возделываемых земель, и если такая тенденция будет продолжаться, то в сельскохозяйственное производство будет вовлекаться всё больше малопригодной для него земли, расходы на это вовлечение и риск деградации окружающей среды при этом будут увеличиваться. Кроме того, население развивающихся стран страдает не столько от недостатков калорий, сколько от недостатка пищи, т.е. от качества пищи – качества энергии пищи.
Около 98% пищи человечество собирает с суши, с океана оно получает 2%, причём преимущественно это животная пища, в то время, как наземное сельское хозяйство даёт примерно в 4 раза больше растительной, чем животной пищи. Вся собираемая пища составляет примерно 1% чистой или 0.5% валовой продукции биосферы. Но надо ещё учесть пищу, поглощаемую сельскохозяйственными животными и домашним скотом. Они поглощают пищи в 5 раз больше, чем всё человечество.
Кстати, следует отметить, что преимущественно используемые виды домашних животных определяют экологию ландшафта конкретных стран, иногда даже в большей степени, чем культура и экономика. Например, в Новой Зеландии экология ландшафта определяется овцами (отношение числа голов домашнего скота к численности населения составляет 43:1, а в среднем по миру 4.7:1, а в Японии 0.6:1), поэтому в Японии экология ландшафта определяется рыбой.
Сейчас плотность населения составляет примерно 1 человек на 2.3 га суши (6 млрд. человек на 14.109 га суши). Если учесть и домашних животных, то получим 1 эквивалент популяции примерно на 0.5 га суши. Если в следующем веке население удвоится и если мы по-прежнему хотим употреблять те же объёмы пищи, то на все нужды каждого консумента массой 50 кг придётся всего 0.25 га.
1.1.6. Пищевые цепи, пищевые сети и трофические уровни
Перенос энергии пищи от его источника – автотрофов (растений) – через ряд организмов, происходящий путём поедания одних организмов другими, называется пищевой цепью.
При каждом очередном переносе большая часть потенциальной энергии (80÷90%) теряется, переходя в тепло. Поэтому чем короче пищевая цепь (чем ближе организм к её началу – солнечной энергии), тем больше количество энергии, доступной для популяции.
Пищевые цепи можно разделить на два основных типа: пастбищная цепь, которая начинается с зелёного растения и идёт далее к пасущимся растительноядным животным и к их хищникам, и детритная цепь, которая от мёртвого органического вещества идёт к микроорганизмам, а затем к детритофагам и к их хищникам. Пищевые цепи не изолированы одна от другой, а тесно переплетаются друг с другом, образуя так называемые пищевые сети.
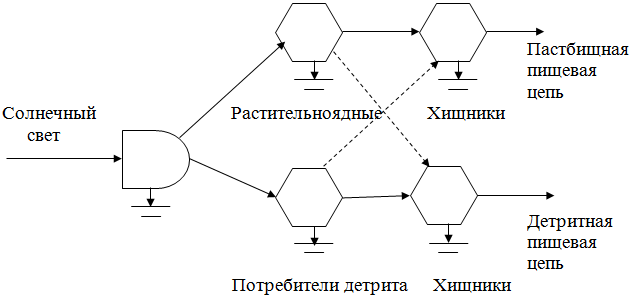
Наиболее упрощенные пастбищная и детритная пищевые цепи объединены в пищевую сеть в виде Y-образной или двухканальной диаграммы потока энергии.
Величины тех частей энергии чистой продукции, которые текут по двум путям, различны в экосистемах разного типа и часто варьируют по сезонам или по годам в одной и той же экосистеме. На некоторых мелководьях и на интенсивно используемых пастбищах и в степях по пастбищной цепи может идти 50% и более чистой продукции. Напротив, прибрежные марши, океаны, леса, да и большинство природных экосистем, функционируют как детритные системы; в них 90% и более процентов автотрофной продукции потребляется гетеротрофами только после того, как листья, стебли и другие части растений отмирают, подвергаются «переработке», превращаясь в диспергированное или растворённое органическое вещество, поступающее в воду, донные осадки и почву. Такое отсроченное потребление увеличивает структурную сложность, а также накопительную и буферную ёмкость экосистем.
Тесная связь пастбищной и детритной пищевых цепей приводит к тому, что при изменении уровня энергетического воздействия извне на экосистему быстро происходит переключение потоков между каналами, что позволяет поддерживать устойчивость экосистем. Не вся пища, съеденная пасущимися животными, усваивается: часть её, например через фекалии, уходит в детритную цепь.
Степень влияния травоядных животных на сообщество зависит не только от количества ассимилированной ими энергии пищи, но и от скорости изъятия живых растений. Прямое изъятие травоядными животными или человеком более 30-50% годового прироста наземной растительности уменьшает способность экосистемы сопротивляться стрессу. Перевыпас скота был одной из причин упадка многих цивилизаций. «Недовыпас » также может оказаться вредным. Если прямое потребление живых растений совершенно отсутствует, то детрит может накапливаться быстрее, чем идёт его разложение микроорганизмами. Это замедляет круговорот минеральных веществ, и, кроме того, система может стать пожароопасной.
В сложных природных сообществах организмы, получающие свою энергию от Солнца через одинаковое число ступеней, считаются принадлежащими к одному трофическому уровню. Так, зелёные растения занимают первый трофический уровень (уровень продуцентов), травоядные – второй (уровень первичных консументов), первичные хищники, поедающие травоядных, – третий (уровень вторичных консументов), а вторичные хищники – четвёртый (уровень третичных консументов). Эта трофическая классификация относится к функциям, а не к видам как таковым. Популяция данного вида может занимать один или несколько трофических уровней, смотря по тому, какие источники энергии она использует. Поток энергии через трофический уровень равен общей ассимиляции (А) на этом уровне, которая в свою очередь равна продукции (Р) биомассы плюс дыхание (R):
A = P + R .
При переносе энергии между трофическими уровнями часть потенциальной энергии теряется. Прежде всего, растение фиксирует лишь малую долю поступающей энергии солнечного света (около 1%). Поэтому число консументов (например, людей), которые могут прожить при данном выходе первичной продукции, сильно зависит от длины пищевой цепи; переход к каждому следующему звену в нашей традиционной сельскохозяйственной пищевой цепи уменьшает доступную энергию примерно на порядок величины (т.е. в 10 раз). Поэтому если в рационе увеличивается содержание мяса, то уменьшается число людей, которых можно прокормить.
Эффективность i-го трофического уровня принято оценивать, как отношение Ai/Ai-1, где Ai – ассимиляция i-ым трофическим уровнем. Для первого (автотрофного) трофического уровня она составляет 1-5%, для последующих – 10-20%.
Может озадачить низкая эффективность природных экосистем в сравнении с высокими КПД электромоторов и других двигателей. Но на самом деле, долгоживущие, крупномасштабные экосистемы нельзя приравнивать в этом отношении к недолговечным механическим системам. Во-первых, в живых системах много «горючего» затрачивается на «ремонт» и самоподдержание, а при расчете КПД двигателей не учитываются амортизация и расходы энергии на ремонт. Во-вторых, в определенных условиях быстрый рост, который повышает потребление энергии, может иметь большее значение для выживания, чем максимальная эффективность использования энергии пищи или горючего.
Для экосистем важно понимать, что любое повышение их эффективности искусственным путём обернется увеличением затрат на ее поддержание. Всегда наступает такой предел, после которого выигрыш от роста эффективности сводится на нет ростом расходов, не говоря уже о том, что система может войти в опасное колебательное состояние, грозящее разрушением. Индустриализованные экосистемы, возможно, уже достигли такой стадии, когда увеличение расходов приводит к все меньшей отдаче.
1.1.7. Концентрация токсичных соединений при продвижении по пищевым цепям
Некоторые вещества по мере продвижения по пищевой цепи не рассеиваются, а, наоборот, накапливаются. Это так называемое концентрирование в пищевой цепи нагляднее всего демонстрируют некоторые устойчивые радионуклиды и пестициды. Например, коэффициент накопления (соотношение количества вещества в тканях и в окружающей среде) радиоактивного фосфора в яйцах гусей равен 2 млн. Таким образом, «безопасные» выбросы в реку могут стать крайне опасными для высших звеньев пищевой цепи.
Изучение детритной пищевой цепи показывает, что благодаря многократному поглощению с начала детритной пищевой цепи в ней должно накапливаться любое вещество, быстро сорбирующееся на частицах детрита и почвы и растворяющееся в кишечнике. Такое накопление подтверждено для ДДТ. У рыб и птиц накоплению способствуют значительные жировые отложения, в которых концентрируется ДДТ. Широкое применение ДДТ приводит к уничтожению целых популяций хищных птиц, таких, как скопы, сапсаны, пеликаны и детритофагов таких, как раков и крабов. Поэтому производство ДДТ в 70-х годах было запрещено.
1.1.8. Качество энергии
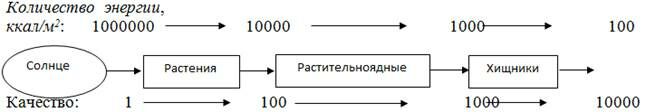
Энергия характеризуется не только количественными, но и качественными параметрами. Одинаковые количества разных форм энергии могут сильно различаться по своему рабочему потенциалу. Высококонцентрированные формы, такие, как энергия нефти, обладают более высоким рабочим потенциалом и соответственно более высоким качеством, чем такие « разбавленные» формы, как солнечный свет, а солнечный свет в свою очередь обладает более высоким качеством по сравнению с еще более рассеянной тепловой энергией.
Качество энергии измеряется количеством данного типа энергии, затрачиваемым на получение другого типа в цепи превращений энергии. По мере того как в этой цепи уменьшается количество энергии, пропорционально на каждом этапе повышается качество той ее доли, которая действительно перешла в новую форму.
Другими словами, при снижении количества повышается качество. Удобным показателем качества может быть количество калорий солнечного света, которое должно рассеяться, чтобы получилась одна калория более высококачественной формы (энергии пищи). Чтобы солнечный свет выполнял работу, производимую нефтью, необходимо повысить его качество в 2000 раз.
Произведение действительного потока энергии на каком-то уровне на показатель качества называется затраченной энергией этого компонента. Доступность энергии ресурса зависит от качества ресурса, например его пищевой ценности.
Метаболизм и размеры особей
Если Вам понравилась эта лекция, то понравится и эта - Новые нетрадиционные религиозные движения и культы.
Метаболизм – это обмен веществ и энергии в биосистеме и его размер значительным образом зависит от размеров особей популяций на определенном трофическом уровне.
Размер урожая биомассы на корню, который может поддерживаться постоянным потоком энергии через пищевую цепь, в значительной степени зависит от размера особей. Чем меньше организм, тем выше его удельный метаболизм (то есть метаболизм в пересчете на 1 г или на 1 кал биомассы) и тем меньше биомасса, которая может поддерживаться на данном трофическом уровне экосистемы. И наоборот, чем крупнее организм, тем выше биомасса на корню. Так, "урожай" бактерий, имеющихся в любой данный момент, будет гораздо ниже "урожая" рыбы или млекопитающих, хотя эти группы, возможно, используют одинаковое количество энергии.
Интенсивность обмена у отдельных организмов и их ассоциаций часто оценивается по скорости потребления кислорода, то есть по дыханию. У животных наблюдается общая тенденция к увеличению интенсивности обмена в расчете на организм пропорционально степени 2/3 роста объёма (или массы), то есть интенсивность метаболизма на грамм биомассы растет с уменьшением длины.
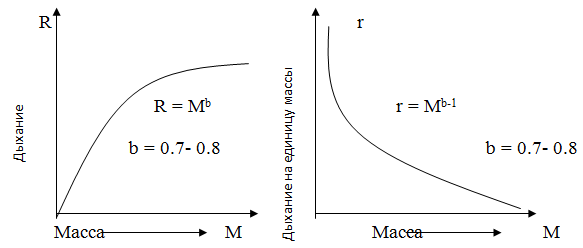
Вторая кривая наглядно демонстрирует рост удельной (по массе) интенсивности метаболизма с уменьшением размеров особь. Большое влияние на данный характер этой зависимости оказывают процессы диффузии через поверхность организмов: действительно, у крупных организмов площадь поверхности, через которую может идти диффузия на грамм массы меньше, чем у мелких животных.
На интенсивность метаболизма влияют и другие факторы. Например, у теплокровных животных интенсивность дыхания выше, чем у холоднокровных животных такого же размера.


















