
Физико-химические процессы, протекающие при брожении
Физико-химические процессы, протекающие при брожении
Анаэробное брожение — это процесс, посредством которого микроорганизмы получают химическую энергию из глюкозы и других субстратов в отсутствии или при дефиците молекулярного кислорода. Его следует рассматривать как простейшую форму биологического механизма, обеспечивающего получение энергии из питательных веществ.

Дрожжевые клетки и молочнокислые бактерии - факультативные анаэробы. Главным клеточным «топливом» при анаэробном получении энергии служат шестиуглеродные сахара, в первую очередь D-глюкоза; при спиртовом брожении, вызываемом дрожжами, пентозы не сбраживаются. Некоторые бактерии способны извлекать энергию, сбраживая пентозы, жирные кислоты или аминокислоты. Среди многих способов сбраживания глюкозы наиболее важную роль играют два тесно связанных между собой — гомоферментативное молочнокислое брожение и спиртовое брожение.
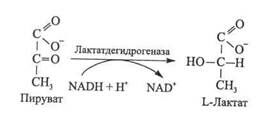
При гомоферментативном молочнокислом брожении молекула гексозы расщепляется на две трехуглеродные молекулы молочной кислоты (лактата), представляющей собой единственный конечный продукт этого процесса.
ФИЗИКО-ХИМИЧЕСКИЕ ПРОЦЕССЫ, ПРОТЕКАЮЩИЕ ПРИ ГЛИКОЛИЗЕ
И СПИРТОВОМ БРОЖЕНИИ
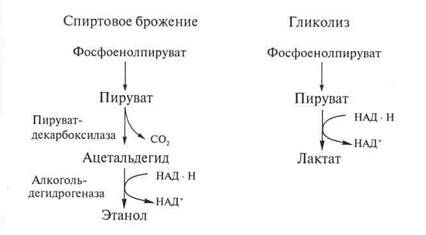
При спиртовом брожении шестиуглеродная молекула глюкозы расщепляется на две двууглеродные молекулы этанола (С2Н5ОН) и две молекулы диоксида углерода (С02). Спиртовое брожение осуществляется тем же ферментативным путем, что и гликолиз, с той разницей, что последняя, завершающая реакция заменена здесь двумя другими, в результате которых трехуглеродные фрагменты разрушаются до этанола и диоксида углерода.
Оба эти процесса включают окислительно-восстановительные реакции. Об этом отчетливо свидетельствуют конечные продукты спиртового брожения; этанол можно рассматривать как относительно восстановленное соединение, т.к. его молекула содержит определенное количество водорода, а диоксид углерода — как относительно окисленное, поскольку водород в его молекуле отсутствует. В конечных продуктах гликолиза окислительно-восстановительный процесс не столь очевиден. Однако один конец молекулы молочной кислоты (метильная группа) находится в более восстановленном состоянии, чем другой ее конец (карбоксильная группа). В молекуле глюкозы атомы водорода распределены более равномерно.
Рекомендуемые материалы
Суммарные уравнения гликолиза и спиртового брожения имеют вид:
|
|

Процесс гликолиза сопровождается сильным уменьшением свободной энергии, составляющим 135,65 кДж. Это необратимый процесс, смещенный в сторону образования лактата.
Гликолиз объединяет три типа химических превращений, пути которых взаимосвязаны: 1) реакции, вызывающие распад глюкозы с образованием конечного продукта - лактата
2) реакции, в результате которых неорганический фосфат (Фн) становится концевой группой АТФ и
3) окислительно-восстановительные реакции - путь переноса электронов.
В гликолиз вовлекаются и другие углеводы — крахмал и гликоген, простые сахара (не D-глюкоза) при участии некоторых дополнительных ферментов. Фермент а-1,4-глюканфосфорилаза катализирует общую реакцию, в которой (глюкоза)n означает полисахаридную цепь глюкана, а (глюкоза) — туже цепь, но укороченную на одну единицу:
(Глюкоза)n +НР04 -2 ↔ (Глюкоза)п_1 + Глюкоза – 1 - фосфат.
Фермент атакует нередуцирующий конец цепи до тех пор, пока не достигнет точки ветвления, т.е. до α-1,6-связи. Конечным продуктом его действия оказывается остаточный декстрин. Для возобновления действия а-1,4-глюканфосфорилазы должен предварительно вступить в действие фермент α - 1,6-глюкозидаза, гидролизу-ющий α -1,6-связь в точке ветвления и открывает для действия α -1,4-глюканфосфорилазы новый участок полисахаридной цепи.
D-манноза и D-фруктоза могут фосфорилироваться в положении 6под действием гексокиназы. D-галактоза включается в гликолиз в результате фосфорилирования, катализируемого галактокиназой в присутствии АТФ. В гликолиз могут вовлекаться и пентозы.
Для организмов, подобных дрожжам, сбраживающим глюкозу до спирта и С02, а не до молочной кислоты, процесс брожения совпадает с гликолизом во всем, за исключением концевого этапа, катализируемого лактатдегидрогеназой. При спиртовом брожении этот процесс заменен двумя другими ферментативными реакциями, катализируемыми соответственно пируватдекарбосилазой и алкоголъдегидрогеназой.
Пируватдекарбоксилаза требует наличия ионов Mg2+ и ко-фермента тиаминпирофосфата (ТПФ) - эфира пирофосфорной кислоты и тиамина (витамина В1). Тиаминпирофосфат служит переносчиком активных альдегидных групп
В заключительной стадии процесс спиртового брожения аце-тальдегид восстанавливается до этанола под действием фермента алкогольдегидрогеназы; при этом роль восстановителя играет HADH + Н+:
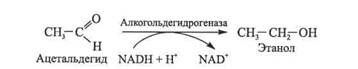
В это уравнение не входят HADH и HAD+, так как HADH, образованный в процессе гликолиза, опять превращается в HAD+ в результате восстановления уксусного альдегида в этанол. Такая регенерация HAD+ обеспечивает в анаэробных условиях непрерывное течение гликолиза.
Реакции, обеспечивающие выделение энергии, сохраняемой в форме АТФ, при гликолизе и при спиртовом брожении идентичны.
Спиртовое брожение является важнейшей стадией в технологии хлеба. Его цель - разрыхление теста диоксидом углерода, придание тесту физических свойств, обеспечивающих его разделку, расстойку и выпечку. Образующиеся при брожении теста под действием дрожжевых клеток наряду с диоксидом углерода и этанолом побочные продукты, обусловливают вкус и аромат хлеба.
Брожение ведется при температуре 27 — 35 °С, величина рН изменяется при брожении от 6,0 - 5,8 до 5,0 и ниже, что связано с растворением в полуфабрикате диоксида углерода и накоплением органических кислот.
Молочнокислое брожение в технологии ржаного хлеба является основной стадией производства. В процессе жизнедеятельности гомо- и гетероферментативных молочнокислых бактерий накапливается достаточные количества молочной кислоты, необходимой для ингибирования а-амилазы, активность которой в ржаной муке весьма высокая. Чрезмерная деятельность этого фермента приводит к интенсивной декстринизации крахмала, особенно на стадии выпечки, когда β-амилаза уже инактивирована.
Именно молочной кислоте принадлежит основная роль в придании ржаному хлебу специфического вкуса и приятного аромата. Кроме того, образующийся в результате жизнедеятельности гетероферментативных молочнокислых бактерий диоксид углерода участвует в разрыхлении заквасок и теста.
В зависимости от соотношения гомо- и гетероферментативных бактерий и от условий их жизнедеятельности в закваске и тесте продукты брожения будут накапливаться в разных количествах. Более высокая активность гетероферментативных бактерий обеспечивает разрыхленность ржаного теста. Однако в тесте при действии этих бактерий накапливается также значительное количество уксусной кислоты. В результате хлеб приобретает неприятный кислый вкус. Превалирующее гомоферментативное брожение способствует накоплению в тесте в достаточном количестве молочной кислоты, придающей хлебу мягкий, приятный кисловатый вкус.
ФИЗИКО-ХИМИЧЕСКИЕ ПРОЦЕССЫ, ПРОТЕКАЮЩИЕ ПРИ ДРУГИХ ТИПАХ БРОЖЕНИЯ
Помимо указанных выше типов брожения существуют и другие типы, многие из которых представляют собой варианты пути Эмбдена — Мейергоффа.
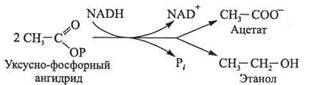
При гетероферментативном молочнокислом брожении из каждой молекулы глюкозы образуется по одной молекуле молочной кислоты, этанола и диоксида углерода. Кроме молочной кислоты, гетероферментативные молочнокислые бактерии образуют уксусную кислоту. Катаболизм глюкозы при этом типе брожения осуществляется до глицеральдегид-3-фосфата по фосфоглюконатному пути. Образовавшийся ангидрид уксусной и фосфорной кислот превращается в уксусную кислоту или восстанавливается в этанол:
 |
При других типах брожения углеводов образуются такие конечные продукты, как пропионовая, масляная, янтарная кислоты и ацетон.
Бактерии рода Clostridium являются возбудителями масляно-кислого брожения, суммарное уравнение которого приведено ниже:
С6Н1206 + 2 ADP + 2 Р, + 2 NADH + 2 Н+ >
СН3-СН2 - СН2 - СООН + 2 NAD + 2 Н2 + 2 С02 + 2 АТР + 2 Н20.
Маслянокислое брожение протекает в средах, куда ограничен доступ кислорода, при наличии субстрата в виде полисахаридов, моносахаридов, органических кислот, спирта. Наряду с масляной кислотой, диоксидом углерода, водородом маслянокислые бактерии могут образовывать в достаточно больших количествах и другие продукты брожения. Так, Clostridium acetobutylicum при сбраживании глюкозы вначале образует масляную кислоту. С подкис-лением среды индуцируются ферменты, в результате деятельности которых накапливаются ацетон и бутанол. Например, синтезированная в бактериальной клетке ацетоацетатдекарбоксилаза в комплексе с тиолазой из ацетил-СоА образуют ацетон:
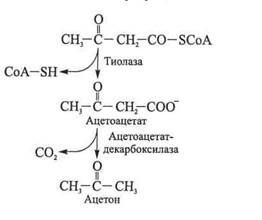
Другим промежуточным продуктом в результате действия фермента Clostridium acetobutylicum является ацетальдегид, образующийся из ацетил-СоА. Последний при участии NADH и ал-когольдегидрогеназы восстанавливается в этанол. Этот тип брожения называют ацетонобутиловым.
Спиртовое, гомоферментативное молочнокислое и масляно-кислое брожения являются основными. Все другие представляют собой комбинацию этих основных. Так, гетероферментативное молочнокислое, пропионовокислое брожения можно представить как комбинированное гомоферментативное молочнокислое и спиртовое; ацетонобутиловое — как маслянокислое и спиртовое; брожение пектиновых веществ, клетчатки - разновидность маслянокислого брожения.
ФИЗИКО-ХИМИЧЕСКИЕ ПРОЦЕССЫ, ПРОТЕКАЮЩИЕ ПРИ МЕТАБОЛИЗМЕ МИКРООРГАНИЗМОВ В ХЛЕБОПЕКАРНЫХ ПОЛУФАБРИКАТАХ
В технологии хлебобулочных изделий для обеспечения процессов спиртового брожения и гликолиза применяют хлебопекарные прессованные дрожжи и молочнокислые бактерии, содержащиеся в бактериальных хлебопекарных заквасках. Микробиологическим процессам, протекающим под их действием в полуфабрикатах, присущи огромные потенциальные возможности.
Отличительной особенностью дрожжевой клетки является— то, что структуры высшего порядка белков и нуклеиновых кислот в ней определяют ее метаболизм. В состав дрожжевой клетки входят биологически важные макромолекулы, сложная организация и воспроизведение которых способствуют ее жизнедеятельности. Следует отметить и тот факт, что все компоненты клетки (белки, нуклеиновые кислоты, липиды, углеводы) непрерывно изменяются, постоянно обновляются.
Метаболизм дрожжевых клеток и бактерий зависит от их способности пропускать воду и растворенные в ней вещества, т.е. зависит от их проницаемости. Эта способность неодинакова у дрожжевых клеток и молочнокислых бактерий. Проблема проницаемости имеет большое значение в механизме обмена веществ, в вопросах распределения веществ между клетками, возникновения биоэлектрических потенциалов.
В основе проникновения вещества в клетку лежит процесс диффузии, математическую трактовку которого дал Фик (1955). По закону Фика, скорость диффузии dm/dt прямо пропорциональна градиенту концентрации dc/dx и площади поверхности s, через которую осуществляется диффузия:
|
|

Коэффициент пропорциональности D в уравнении (6.6) носит название константы диффузии.
Скорость проникновения вещества в клетку определяется по формуле, предложенной Коллендером и Берлундом (Collander, Barlund):

Вычисление константы проницаемости проводят по формуле (6.7), которую можно представить в следующем виде:
Принимая т/с, равным v (v — объем клетки), и интегрируя, получаем:
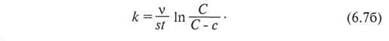
Считая v и s постоянными в течение всего времени эксперимента, получим:
|
|

Проникновение вещества может происходить и в направлении, обратном концентрационному градиенту, при наличии другого градиента, противоположно направленного, например, электрического.
При исследовании проницаемости следует постоянно учитывать роль метаболических процессов и в обеспечении концентрационного градиента между клеткой и средой. Так, включение поступающего в клетку аниона фосфора в молекулы нуклеотидов, нуклеиновых кислот, нуклеопротеидов и фосфолипидов приводит к снижению концентрации неорганического фосфора в клетке и тем самым поддерживает градиент этого аниона между клеткой и средой.
Важную роль в накоплении веществ в клетке играет и химическая адсорбция.
Направление потока веществ не остается раз навсегда постоянным: оно может измениться на обратное при нарушении соотношений между процессами синтеза и распада важнейших высокомолекулярных компонентов протоплазмы. Так, в молодых клетках происходит накопление ионов калия и фосфатов за счет включения последних в молекулы АТФ. Напротив, по мере старения клеток происходит распад органических фосфатов с отщеплением и последующим выходом фосфатов, а также ионов калия в среду. Иначе говоря, направление потока веществ изменяется на противоположное.
При всем разнообразии строения и физико-химических свойств молекул веществ существуют в основном два пути их проникновения в клетку: 1) через субмикроскопические заполненные водой поры, пронизывающие протоплазму и соединяющие ее с внешней средой, проникают молекулы водорастворимых веществ и ионы; 2) за счет растворения в липидах, входящих в состав протоплазмы и особенно ее поверхностных слоев, проникают водонера-створимые органические соединения.
Проницаемость клеток для органических молекул уменьшается по мере возрастания количества гидроксильных, карбоксильных и аминогрупп.
Для диффузии молекул водорастворимых соединений, при условии если их радиус не превышает радиуса гексоз, применимо уравнение Товерта:
Джекобс (М. Jacobs), исходя из современной теории электрического строения молекул, показал, что все химические соединения, а также отдельные, входящие в их состав радикалы, можно разбить на две большие группы:
1) электрически симметричные, т.е. те, у которых центры положительных и отрицательных зарядов совпадают друг с другом, названные неполярными, или гомеополярными (метиловые, этиловые, фениловые группы);
2) электрически несимметричные, центры положительных и отрицательных зарядов которых не совпадают, - полярные или гетерополярные (соединения с группами СООН, ОН и NH2).
Неполярные соединения хорошо растворимы в липидах, хлороформе и ацетоне, имеют низкую диэлектрическую постоянную, а полярные хорошо растворимы в воде и характеризуются высокой диэлектрической постоянной.
Исключение из этого правила составляет глюкоза, плохо растворимая в липидах, но легко проникающая в клетку. Возможно, что глюкоза нарушает глобулярную структуру некоторых белков. Аномально высокая проницаемость клеток для глицерина может быть отнесена на счет его способности образовывать димеры, в которых маскирующие друг друга полярные гидроксильные группы соседних молекул не препятствуют липидорастворимости.
Поступление веществ из внешней среды обеспечивают клеточная стенка дрожжевой клетки, цитоплазматическая мембрана, пиноцитоз и эндоплазматическая сеть.
Бактерии имеют относительно простое строение. Бактериальная клетка состоит из протоплазмы (цитоплазмы) с различными включениями, которая от внешней среды отделена плотной оболочкой — клеточной стенкой. Стенки бактериальных клеток представляют собой жесткие, пористые, замкнутые структуры, обеспечивающие клеткам защиту от физических воздействий, препятствующие набуханию и разрыву клеточной мембраны. Клеточные стенки бактерий принимают участие во многих физиологических и физико-химических процессах, выполняя различные функции в жизни клетки. Оболочки клеток химически активны, в них происходит множество химических реакций. Цитоплазматическая мембрана бактерий контролирует поступление питательных веществ и выход продуктов метаболизма из клеток.
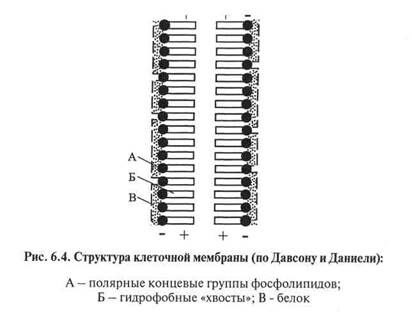
Физико-химические процессы проницаемости дрожжевых клеток для неполярных соединений объясняются липидной структурой клеточной мембраны. Липид ведет себя как растворитель для неполярных участков белков, стероидов и т.д. В цитоплаз-матической мембране белковые молекулы, расположенные между слоями липидов, могут изменять свое положение. Липиды, согласно глобулярной структуре биомембраны дрожжей, организованы в биослои, но они не являются непрерывными (рис. 6.4).
Толщину биомолекулярного слоя можно вычислить по формуле Фрикке:
|
|
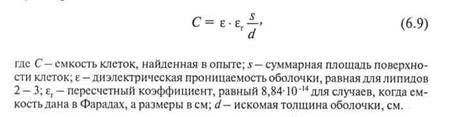
Расчеты, произведенные на основании опытных данных, показали, что толщина клеточной мембраны приблизительно равна 32 А, что соответствует бимолекулярному слою жирных кислот с длиной углеводородной цепочки в 16 - 18 углеродных атомов.
Обратите внимание на лекцию "Объектно-ориентированные CASE-средства".
В клетку могут проникать вещества, нерастворимые в липидах, в первую очередь, минеральные ионы, сахара и аминокислоты.
Для того чтобы молекула или ион водорастворимого вещества проникли в клетку, они должны обладать некоторым добавочным количеством энергии. Пересекая клеточную мембрану, они должны, во-первых, оторваться от окружающей их сольватной оболочки, во-вторых, раздвинуть плотно расположенные молекулы поверхностного слоя на границе клетка — омывающий ее раствор, в-третьих, преодолеть взаимодействие своих полярных групп с полярными группами, расположенными внутри канала поры, в-четвертых, преодолеть энергетический барьер на границе поверхностной мембраны и водного раствора коллоидов и ионно-дисперсных веществ протоплазмы.
Изменяя активную реакцию внешней среды, можно увеличить или уменьшить количество недиссоциированных молекул и тем усилить или ослабить проникновение вещества в клетку.
Процессы обмена веществ в дрожжевой клетке протекают с участием биологических катализаторов, в состав которых входят экзо- и эндоферменты. Взаимосвязь дрожжевых клеток с окружающей средой, приспособление к ней создают благоприятные физико-химические условия и предопределяют активность их ферментов, ускоряют и обеспечивают внутриклеточные биохимические превращения.
В условиях хлебопекарного производства факторами, влияющими на рациональный ход технологического процесса и качество продукции, являются исходная биологическая активность дрожжевых клеток и способность их усваивать питательные вещества. Эффективность их усвоения зависит от ряда физико-химических факторов: проницаемости клеточной мембраны, возникновения биоэлектрических потенциалов, величин концентрационного и электрического градиентов, наличия водо- и нерастворимых веществ в окружающей среде, рН среды, величины энергетического барьера, активности и др. Уникальным свойством дрожжевой клетки является непрерывное получение энергии - химической, электрической, необходимой для ее метаболизма и физиологических функций. Энергия в клетке накапливается в результате окислительно-восстановительных реакций.
Используя научные концепции метаболизма дрожжей, коэффициент распределения питательных веществ между клетками и окружающей средой, можно регулировать их активность при брожении хлебопекарных полуфабрикатов.



















