
Часть 1
БИОХИМИЯ НЕРВНОЙ ТКАНИ
По своему соcтаву и процессам метаболизма нервная ткань значительно отличается от других тканей.
Центральная функциональная клетка нервной ткани - нейрон - связана с помощью дендритов и аксонов с такими же клетками и клетками других типов, например, с секреторными и мышечными клетками. Клетки разделены синаптическими щелями. Связь между клетками осуществляется путем передачи сигнала. Сигнал проходит от тела нейрона по аксону до синапса. В синаптическую щель выделяется вещество-медиатор. Медиатор вступает в связь с рецепторами на другой стороне синаптической щели. Это обеспечивает восприятие сигнала и генерацию нового сигнала в клетке-акцепторе.
ФУНКЦИИ НЕРВНОЙ ТКАНИ
1. Генерация электрического сигнала (нервного импульса)
2. Проведение нервного импульса
3. Запоминание и хранение информации.
4. Формирование эмоций и поведения.
5. Мышление.
Рекомендуемые материалы
ОСОБЕННОСТИ ХИМИЧЕСКОГО СОСТАВА И МЕТАБОЛИЗМА НЕРВНОЙ ТКАНИ
Специфику нервной ткани определяет гематоэнцефалический барьер (ГЭБ). ГЭБ имеет избирательную проницаемость для различных метаболитов, а также способствует накоплению некоторых веществ в нервной ткани. Например, в нервной ткани на долю глутамата и аспартата приходится примерно 70-75% от общего количества аминокислот. Таким образом, внутренняя среда нервной ткани намного отличается по химическому составу от других тканей.
ЛИПИДЫ НЕРВНОЙ ТКАНИ.
ФУНКЦИИ ЛИПИДОВ НЕРВНОЙ ТКАНИ.
1. Структурная: входят в состав клеточных мембран нейронов.
2. Функция диэлектриков (обеспечивают надежную электрическую изоляцию).
3. Защитная. Ганглиозиды являются очень активными антиоксидантами - ингибиторами перекисного окисления липидов (ПОЛ). При повреждении ткани мозга ганглиозиды способствуют ее заживлению.
4. Регуляторная. Фосфатидилинозиты являются предшественниками биологически активных веществ.
Большая часть липидов нервной ткани находится в составе плазматических и субклеточных мембран нейронов и в миелиновых оболочках. В нервной ткани по сравнению с другими тканями организма содержание липидов очень высокое.
Особенность липидного состава нервной ткани: есть фосфолипиды (ФЛ), гликолипиды (ГЛ) и холестерин (ХС), нет нейтральных жиров. Эфиры холестерина можно встретить только в участках активной миелинизации. Сам холестерин синтезируется интенсивно только в развивающемся мозге. В мозге взрослого человека низка активность ОМГ-КоА-редуктазы - ключевого фермента синтеза холестерина. Содержание свободных жирных кислот в мозге очень низкое.
Некоторые нейромедиаторы после взаимодействия со специфическими рецепторами изменяют свою конформацию и изменяют конформацию фермента фосфолипазы С, которая катализирует расщепление связи в фосфатидилинозите между глицерином и остатком фосфата, в результате чего образуется фосфоинозитол и диацилглицерин. Эти вещества являются регуляторами внутриклеточного метаболизма. Диацилглицерин активирует протеинкиназу С, а фосфоинозитол вызывает повышение концентрации Са2+. Ионы кальция влияют на активность внутриклеточных ферментов и участвуют в работе сократительных элементов нервных клеток: микрофиламентов, что обеспечивает передвижение различных веществ в теле нервной клетки, аксоне и растущем кончике аксона. Протеинкиназа С участвует в реакциях фосфорилирования белков внутри нервных клеток. Если это белки-ферменты, то меняется их активность, если это рибосомальные или ядерные белки, то изменяется скорость биосинтеза белков.
Липиды постоянно обновляются. Скорость их обновления различна, но в целом низка.. Некоторые липиды (например: холестерин, цереброзиды, фосфатидилэтаноламины, сфингомиелины) обмениваются медленно - в течение месяцев и даже лет. Исключение составляют фосфатидилхолин и, особенно, фосфатидилинозиты (содержат глицерин, фосфат, спирт (инозит), жирные кислоты) - они обмениваются очень быстро (сутки, недели).
Синтез цереброзидов и ганглиозидов протекает с большой скоростью в развивающемся мозге в период миелинизации. У взрослых почти все цереброзиды (до 90%) находятся в миелиновых оболочках, а ганглиозиды - в нейронах.
НУКЛЕИНОВЫЕ КИСЛОТЫ
Нервные клетки не делятся, значит, не происходит синтез ДНК. Однако, содержание РНК в них самое высокое по сравнению с клетками остальных тканей организма. Скорость синтеза РНК тоже очень велика.
В клетках нервной ткани не могут синтезироваться пиримидины (в нервной ткани отсутствует фермент карбамоилфосфатсинтаза). Пиримидины обязательно должны поступать из крови - ГЭБ для них проницаем. ГЭБ легко проницаем и для пуриновых мононуклеотидов, но, в отличие от пиримидиновых, они могут синтезироваться в нервной ткани.
В нервной ткани, так же, как и в других, нуклеиновые кислоты обеспечивают хранение и передачу генетической информации и ее реализацию при синтезе клеточных белков.
Например, сильные раздражители: громкие звуки, сильные зрительные стимулы и эмоции приводят к повышению скорости синтеза и РНК, и белка в определенных участках мозга. Это указывает на то, что изменения в нервной системе, отражающие индивидуальный опыт организма, кодируются в виде синтезированных макромолекул.
Информация, благодаря которой нейроны устанавливают только определенные связи с определенными нейронами, кодируется в структуре полисахаридных веточек мембранных гликопротеинов. Образование таких связей, не заложенных в период эмбрионального развития, является результатом опыта индивидуального организма и составляет материальную основу для хранения информации, определяющей особенности поведения данного организма.
МЕТАБОЛИЗМ УГЛЕВОДОВ И ОСОБЕННОСТИ ЭНЕРГЕТИЧЕСКОГО ОБЕСПЕЧЕНИЯ НЕРВНОЙ ТКАНИ.
В нервной ткани, составляющей только 2% от массы тела человека, потребляется 20% кислорода, поступающего в организм. При этом энергетические возможности нервной ткани ограничены.
1. Основной путь получения энергии - только аэробный распад глюкозы по ГБФ-пути. Глюкоза является почти единственным энергетическим субстратом, поступающим в нервную ткань, который может быть использован ее клетками для образования АТФ.
2. Проникновение глюкозы в ткань мозга не зависит от действия инсулина, который не проникает через гематоэнцефалический барьер. Влияние инсулина проявляется лишь в периферических нервах.
3. Постоянный и непрерывный приток глюкозы и кислорода из кровеносного русла является необходимым условием энергетического обеспечения нервных клеток. Жесткая зависимость от поступления глюкозы обусловлена тем, что содержание гликогена в нервной ткани ничтожно (0.1% от массы мозга) и не может обеспечить мозг энергией даже на короткое время. С другой стороны, окисления неуглеводных субстратов с целью получения энергии не происходит. Поэтому при гипогликемии и/или даже кратковременной гипоксии в нервной ткани образуется мало АТФ. Следствием этого являются быстрое наступление коматозного состояния и необратимых изменений в ткани мозга.
4. Высокая скорость потребления глюкозы нервными клетками обеспечивается, в первую очередь, работой высокоактивной гексокиназы мозга. В отличие от других тканей, здесь гексокиназа не является ключевым ферментом всех путей метаболизма глюкозы. Гексокиназа мозга отличается низким значением Км и высокой Vmax, обладает в 20 раз большей активностью, чем соответствующий изофермент печени и мышц. Ключевыми ферментами ГБФ-пути в нервной ткани являются фосфофруктокиназа и изоцитратдегидрогеназа. Фосфофруктокиназу ингибируют фруктозо-1,6-бисфосфат, АТФ и цитрат, активируют фруктозо-6-фосфат, АДФ, АМФ и неорганический фосфат. Активность изоцитратДГ даже при нормальном уровне утилизации глюкозы в состоянии покоя максимальна. Поэтому при повышенном энергопотреблении нет возможностей ускорения реакций ЦТК.
5. Образование НАДФН2, который используется в нервной ткани в основном для синтеза жирных кислот и стероидов, обеспечивается сравнительно высокой скоростью протекания ГМФ-пути распада глюкозы.
Энергия АТФ в нервной ткани используется неравномерно во времени.
Так же, как и скелетные мышцы, функционирование нервной ткани сопровождается резкими перепадами в потреблении энергии. Резкое повышение энергозатрат происходит при очень быстром переходе от сна к бодрствованию. Поэтому существует еще одна особенность:
6. Образование креатинфосфата. Он обладает способностью удерживать макроэргические связи:
|
|
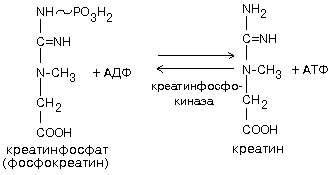
Эта реакция полностью обратима, ее направление зависит от соотношения АТФ/АДФ в клетках нервной ткани. Во время сна накапливается фосфокреатин. Переход к бодрствованию приводит к резкому уменьшению концентрации АТФ - равновесие реакции сдвигается влево, то есть образуется АТФ.
МЕТАБОЛИЗМ АМИНОКИСЛОТ И БЕЛКОВ
Ткань мозга интенсивно обменивается аминокислотами с кровью. Для этого существует специальные транспортные системы: две для незаряженных и еще несколько - для аминокислот, заряженных положительно и отрицательно.
До 75% от общего количества аминокислот нервной ткани составляют аспартат, глутамат, а также продукты их превращений или вещества, синтезированные с их участием (глутамин, ацетильные производные, глутатион, ГАМК и другие). Их концентрации, и, в первую очередь, концентрация глутамата, в нервной ткани очень высоки. Например, концентрация глутаминовой кислоты может достигать 10ммоль/л.
Функции глутамата в нервной ткани:
1. Энергетическая. Глутаминовая кислота связана большим числом реакций с промежуточными метаболитами ЦТК.
2. Глутамат (вместе с аспартатом) принимает участие в реакциях дезаминирования других аминокислот и временном обезвреживании аммиака.
3. Из глутамата образуется нейромедиатор ГАМК.
4. Глутамат принимает участие в синтезе глутатиона - одного из компонентов антиоксидантной системы организма.
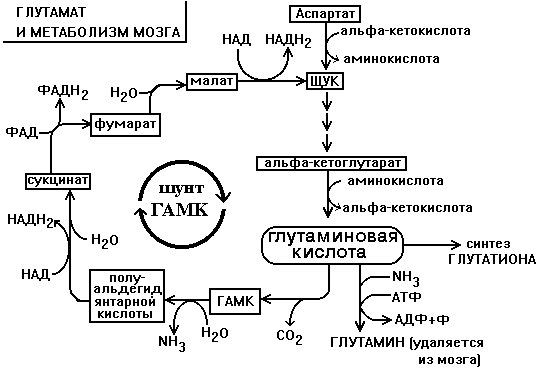
Глутаминовая кислота по праву занимает центральное место в обмене аминокислот мозга. Она используется для образования глутатиона, глутамина и гамма-аминомасляной кислоты. Образуется глутамат из своего кетоаналога - альфа-кетоглутаровой кислоты в ходе реакции трансаминирования. Реакция превращения альфа-КГ в глутамат протекает в ткани мозга с большой скоростью. Образующийся при этом глутамат являетя для ЦТК побочным продуктом. Большое расходование альфа-КГ восполняется за счет превращения аспарагиновой кислоты в метаболит ЦТК - ЩУК (см. рисунок).
Образующаяся из глутамата ГАМК в результате нескольких реакций может быть превращена снова в ЩУК. Так образуется ГАМК-шунт, имеющийся в тканях головного и спинного мозга. Поэтому в этих тканях содержание ГАМК, как промежуточного метаболита циклического процесса, значительно выше, чем в остальных. На образование ГАМК здесь используется до 20% от общего количества глутамата.
Остальные пути метаболизма аминокислот сходны с имеющимися в других тканях.
До сих пор непонятным остается наличие в мозге почти полного набора ферментов орнитинового цикла, не содержащего карбамоилфосфатсинтазы, из-за чего мочевина здесь не образуется.
Ткань мозга способна синтезировать заменимые аминокислоты, как и другие ткани.
НЕЙРОМЕДИАТОРЫ
Нейромедиаторы - это вещества, которые характеризуются следующими признаками:
1. Накапливаются в пресинаптической структуре в достаточной концентрации;
2. Освобождаются при передаче импульса
3. Вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса.
4. Имеют систему для инактивации или транпортную систему для удаления из синапса, обладающие к ним высоким сродством.
Таким образом, нейромедиаторы играют важную роль в функционировании нервной ткани, обеспечивая синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление - в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов.
Рекомендуем посмотреть лекцию "Лекция 12".
ХИМИЧЕСКАЯ КЛАССИФИКАЦИЯ НЕЙРОМЕДИАТОРОВ.
1) Аминокислоты (и их производные). К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
ТАУРИН.
Таурин образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты:
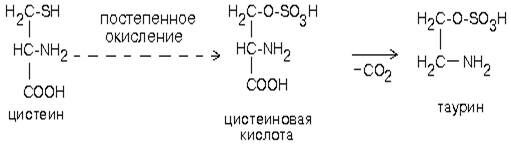
Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.




















