
Гаметогенез и эмбриогенез: дробление, бластула, гаструляция
Тема 3
Гаметогенез. Эмбриогенез
1. Дробление. Типы дробления.
2. Бластула. Типы бластул. Влияние факторов внешней среды на процессы дробления.
3. Гаструляция.
4. Гистогенез и органогенез.
5. Теория зародышевых листков.
6. Производные зародышевых листков.
ИСПОЛЬЗОВАННАЯ И РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Рекомендуемые материалы
Антипчук, Ю.П. Гистология с основами эмбриологии / Ю.П. Антипчук. – М.: Просвещение, 1983. – 240 с.
Алмазов, И.В., Сутулов Л.С. Атлас по гистологии и эмбриологии / И.В. Алмазов, Л.С. Сутулов. – М.: Медицина, 1978. – 148 с.
Гистология / под ред. Ю.И. Афанасьева. – М: Медицина, 1989. – 361 с.
Рябов, К.П. Гистология с основами эмбриологии / К.П. Рябов. – Мн.: Высш. шк., 1991. – 289 с.
Биологический энциклопкдический словарь / под ред. М.С. Гилярова. – М.: Сов. Энцикл., 1989. – 864 с.
Практикум по гистологии, цитологии и эмбриологии / под ред. Н.А. Юриной, А.И. Радостиной. – М.: Высш. шк., 1989. – 154 с.
Хэм А., Кормик Д. Гистология / А. Хэм, Д. Кормик. – М.: Мир, 1983. – 192
1. ДРОБЛЕНИЕ
После оплодотворения яйцеклетка начинает делиться. Процесс деления яйцеклетки, приводящий к увеличению числа клеток без их роста, называют дроблением. Характер дробления определяется количеством желтка в яйцеклетке и его распределением. В зависимости от этого дробление может быть полным и частичным.. Дробление, при котором яйцеклетка делится полностью, называют голобластическими, а неполностью, частично – меробластическими.
Полное дробление яйцеклетки может быть равномерным и неравномерным.
Полное равномерное дробление свойственно яйцеклеткам ланцетника, яйца которого имеют мало желтка. При этом виде дробления образующиеся клетки, или бластомеры, имеют примерно одинаковые размеры.
Полное неравномерное дробление свойственно яйцеклеткам хрящевых рыб, земноводных, яйца которых имеют больше желтка. При этом виде дробления образуются неодинаковые по величине бластомеры, причем первые два дробления дают равные бластомеры, а затем бластомеры, расположенные на анимальном полюсе, делятся быстрее, становятся мельче, нежели на вегетативном.
Неполное дробление яйцеклеток может быть поверхностным и дискоидальным.
При неполном поверхностном дроблении, которое присуще многим членистоногим, дробится лишь поверхностная часть яйцеклетки, а центральная ее часть, богатая желтком, не делится.
При неполном дискоидальном дроблении, которое свойственно, например, пресмыкающимся, птицам, дробится лишь небольшой участок поверхности яйцеклетки, где мало желтка, а остальная ее часть, богатая желтком, не делится. В связи с тем что при этом виде дробления поверхностный участок дробящегося яйца имеет форму диска, то и дробление получило наименование дискоидального (рис. 1).
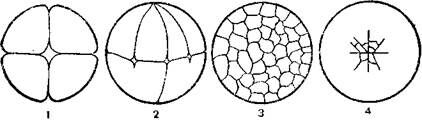 |
Рис. 1. Типы дробления яйцеклеток (ориг.):
1 – полное равномерное; 2 – полное неравномерное; 3 – неполное поверхностное; 4 – неполное дискондальное.
Характер дробления определяется не только количеством желтка и распределением его в яйцеклетке, но и взаимным расположением клеток, которые образуются в результате дробления.
По направлению дробления различают радиальное, спиральное, двусимметричное, или билатеральное, дробление.
При радиальном дроблении, которое свойственно кишечнополостным, иглокожим и многим низшим хордовым, верхний ряд бластомеров располагается точно над нижним.
При спиральном дроблении, которое наблюдается у большинства червей и моллюсков, верхний ряд бластомеров располагается между бластомерами нижнего ряда.
При билатеральном дроблении, которое свойственно круглым червям, асцидиям, бластомеры располагаются симметрично по бокам исходного бластомера. Следует заметить, что в процессе дробления яйцеклетки зачастую имеет место сочетание различных типов дробления.
2. БЛАСТУЛА, ИЛИ ОДНОСЛОЙНЫЙ ЗАРОДЫШ
В процессе дробления возникает многоклеточный однослойный зародыш, который внешне напоминает шар, во многих случаях с полостью в середине. Такой зародыш называют бластулой. Слой клеток, образующий стенку бластулы, называют бластодермой (derma – кожа), а полость бластулы – бластоцелем (koilon – полость) или первичной полостью тела. Когда же при дроблении возникает шаровидный зародыш без полости внутри, похожий на ягоду шелковицы, то его называют морулой (morula ягода шелковицы). Такая бластула свойственна, например, плацентарным млекопитающим.
В зависимости от типа дробления яйцеклетки различают, кроме морулы, еще 5 типов бластул: целобластулу, амфибластулу, стерробластулу, перибластулу и дискобластулу (рис. 2).
Бластулу, бластодерма которой образована одним слоем клеток, а бластоцель занимает центральное положение, называют типичной бластулой или целобластулой. Такая бластула свойственна, например, ланцетнику.
Амфибластула отличается от типичной бластулы тем, что ее бластоцель смещен к анимальному полюсу – зоне мелких бластомеров, а бластодерма представлена несколькими рядами клеток. Такая бластула свойственна земноводным, от которых и получила свое наименование.
Стерробластула (sterros – плотный) состоит из крупных клеток бластодермы, которые настолько глубоко заходят в полость бластоцеля, что от последнего ничего не остается. Такую бластулу имеют некоторые членистоногие.
В перибластуле, возникающей при поверхностном дроблении яйца, клетки бластодермы располагаются по периферии недробящегося желтка и бластоцеля нет. Такой тип бластулы присущ некоторым насекомым.
Дискоидальное дробление ведет к формированию дискобластулы, бластоцель которой имеет вид узкой щели, расположенной под клетками бластодермы, именуемой в данном случае зародышевым диском. Такой тип бластулы наблюдается у костистых рыб, пресмыкающихся, птиц.
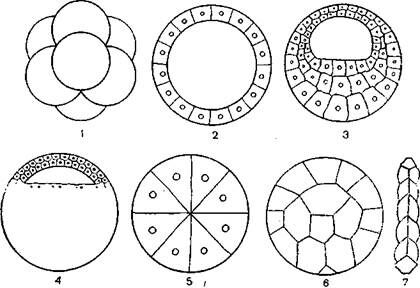
Рис. 2. Типы бластул (ориг.):
/ – морула; 2 – цслобластула; 3 – амфнбластула; 4 – дискобластула; 5 – стерробластула; 6 – перибластула; 7 – плакула.
Рассмотренное многообразие бластул показывает, что на этой стадии развития организмов имеет место дифференцировка клеточного материала зародыша, связанная с количеством и расположением желтка, с содержанием и синтезом ДНК, с разделением веществ в цитоплазме клеток.
На процессы дробления и ранние стадии развития организмов влияют условия внешней среды, поскольку развитие организма происходит в среде, в которой проходило его формирование в историческом развитии. Незначительные отклонения условий среды от типичных сказываются в основном лишь на темпах дробления.
К факторам внешней среды, которые влияют на дробление яйцеклетки и ранние стадии развития организма, следует в первую очередь отнести наличие влаги, химический состав и реакцию среды (рН), световой поток, температуру, наличие питательных веществ, кислорода и др.
3. ГАСТРУЛЯЦИЯ
Гаструляция – это процесс образования двуслойного зародыша.
В более широком представлении гаструляция – сложный процесс химических и морфологических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки, источники зачатков тканей и органов, комплексы осевых органов.
Гаструляция у большинства животных происходит в два этапа. На первом этапе образуются 2 зародышевых листка (эктодерма и энтодерма), а на втором формируется третий 3 зародышевый листок (мезодерма) и происходят дальнейшее формирование эмбриона.
Различают четыре типа, или способа, образования двуслойного зародыша: вселение, или иммиграция; впячивание, или инвагинация; обрастание, или эпиболия; расслоение, или деляминация.
Исходной в процессе филогенетического развития и наиболее примитивной формой гаструляции является вселение, или иммиграция. Такой способ образования двуслойного зародыша состоит в том, что отдельные клетки или группы клеток активно перемещаются из бластодермы в бластоцель. Эти клетки, возникающие вследствие деления клеток бластодермы, оседают на ее внутренней поверхности, образуя внутренний слой клеток, или энтодерму, а клетки бластодермы, расположенные снаружи, именуются уже эктодермой. Если вселение клеток в бластоцель имеет место лишь со стороны одного вегетативного полюса бластулы, то такая иммиграция носит название униполярной или однополюсной, а когда из различных участков бластодермы, – мультиполярной или многополюсной. Униполярная иммиграция свойственна некоторым гидроидным полипам, медузам и почти всем гидромедузам. Мультиполярная иммиграция является редким явлением и наблюдается у некоторых медуз. При иммиграции гастральная полость, или гастроцель (полость первичной кишки), образуется либо одновременно с возникновением двуслойного зародыша, как это имеет место, например, у гидромедуз, либо значительно позже, как у многих видов кишечнополостных. У последних наблюдается столь массовое вселение клеток, что бластоцель ими полностью заполняется, полость исчезает и лишь впоследствии, с обособлением энтодермы, возникает гастроцель, естественно, не имеющий связи с внешней средой. Этот способ гаструляции особенно распространен среди двуслойных животных – кишечнополостных. Все иные способы гаструляции являются производными иммиграции.
Если гаструляция осуществляется путем впячивания, или инвагинации (in— проникновение, vagina – оболочка, влагалище), то однослойная стенка бластулы – бластодерма – прогибается внутрь бластоцеля и достигает противоположной стенки. Впячивание начинается с вегетативного полюса, который распознается по более крупным клеткам, и идет в направлении анимального полюса. По окончании инвагинации зародыш становится двуслойным, поскольку бластодерма образует два слоя клеток: наружный – эктодерму и внутренний – энтодерму. После впячивания формируется полость, которая окружена клетками энтодермы. Эту полость называют гастральной полостью. Гастроцель сообщается с внешней средой отверстием, которое называют первичным ртом или бластопором). Края этого отверстия именуются губами.
В процессе эволюционного развития представителей животного царства формирование бластопора при гаструляции послужило основанием для разделения животных на две большие группы по такому признаку, как образование первичного рта. У червей, моллюсков, членистоногих первичный рот, развиваясь и дифференцируясь, превращается в постоянный, или дефинитивный, рот взрослой особи. Таких животных называют первичноротыми. У иглокожих, кишечнодышащих, хордовых бластопор превращается в заднепроходное, или анальное, отверстие, или в нервно-кишечный канал, который находится на заднем конце эмбриона, а ротовое отверстие возникает заново на переднем конце брюшной поверхности зародыша. Таких животных именуют вторичноротыми. Гаструляция путем впячивания очень распространена у низших вторичноротых и хордовых.
Судьба бластоцеля при инвагинации может быть двоякой: он может сохраниться в том случае, если клеточные слои эктодермы и энтодермы не соприкасаются между собой и тогда у зародыша наблюдаются одновременно две полости – бластоцель и гастроцель. Если же клеточные слои эктодермы и энтодермы при впячивании соприкасаются на всем протяжении, то бластоцель исчезает и у зародыша наблюдается лишь одна полость – полость первичной кишки, или гастроцель.
У некоторых позвоночных животных в связи с увеличением количества желтка в яйцеклетке и незначительными размерами бластоцеля, оттесняемого к анимальному полюсу, гаструляция лишь по типу инвагинации становится невозможной. Тогда у некоторых круглоротых и земноводных к впячиванию присоединяется обрастание, или эпиболия. Гаструляция путем эпиболии состоит в том, что клетки бластодермы анимального полюса бластулы делятся значительно быстрее, чем клетки вегетативного полюса. В связи с этим мелкие клетки анимального полюса постепенно снаружи наползают на крупные клетки вегетативного полюса, обрастая их и образуя эктодерму, а клетки вегетативного полюса формируют энтодерму. При гаструляции путем эпиболии в чистом виде, как это имеет место у некоторых беспозвоночных, вначале нет ни бластопора, ни гастроцеля, которые возникают значительно позже. В тех же случаях, когда эпиболия сопутствует инвагинации, как это наблюдается у некоторых костистых рыб, земноводных, то возникает бластопор, а также полость первичной кишки, которые существуют наряду с бластоцелем.
У костистых рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих, яйцеклетки которых имеют очень много желтка, в первой фазе гаструляции двуслойный зародыш образуется путем расслоения, или деляминации, клеточного слоя бластодермы. Этот тип образования двуслойного зародыша наблюдается у кишечнополостных, а также у высших плацентарных млекопитающих, яйцеклетки которых утратили большие запасы желтка. Гаструляция путем деляминации состоит в делении клеток бластодермы, причем возникающие при этом дочерние клетки оттесняются в бластоцель и образуют внутренний слой клеток, или энтодерму, а наружный слой клеток, или прежняя бластодерма, преобразуется в эктодерму. Когда происходит оттеснение дочерних клеток в бластоцель, мы имеем дело с так называемой первичной деляминацией, а при вторичной деляминации клетки будущей эктодермы, наоборот, оттесняются кнаружи. При этом способе гаструляции бластопор не образуется и гастроцель, естественно, не сообщается с внешней средой.
Следует отметить, что, несмотря на своеобразность отдельных способов гаструляции, в процессе образования многослойного зародыша зачастую наблюдается комбинация различных типов гаструляции – двух и даже трех типов.
Образование третьего слоя клеток, или третьего зародышевого листка – мезодермы, в процессе эмбрионального развития животных осуществляется четырьмя способами: телобластическим, энтероцельным, эктодермальным и смешанным, или переходным.
У первичноротых животных образование мезодермы осуществляется за счет нескольких крупных клеток, располагающихся во время гаструляции на границе между эктодермой и энтодермой по бокам бластопора. Происхождение этих клеток не выяснено, равно как и отношение их к какому-либо из зародышевых листков, поскольку они обособляются уже на стадии дробления. Дочерние клетки указанных клеток располагаются между эктодермой и энтодермой и формируют третий слой клеток – мезодерму. Поскольку исходные клетки, образующие мезодерму, в течение всего времени располагаются на заднем конце зародыша, то соответственно своей локализации они получили название телобластов, а сам способ образования мезодермы – телобластического. Вторичная полость тела зародыша, или целом, образуется клетками мезодермы вследствие их расслоения.
У вторичноротых, включая низших хордовых, мезодерма образуется из клеток энтодермы. При этом способе образования мезодермы клетки стенки первичной кишки, или энтодермы, интенсивно размножаясь, образуют по бокам первичной кишки мешковидные выпячивания в бластоцель. Эти выпячивания, врастая в бластоцель, располагаются между эктодермой и энтодермой. В дальнейшем энтодермальные выпячивания отшнуровываются от энтодермы, а целостность стенки первичной кишки и энтодермальных выростов восстанавливается путем размножения клеток. После обособления от первичной кишки клеточный материал энтодермального происхождения именуется третьим зародышевым листком или мезодермой. Разрастаясь, мезодерма заполняет всю полость бластоцеля, а полость мешкоподобных выростов, являясь по происхождению полостью гастроцеля, именуется вторичной полостью тела. Этот способ образования мезодермы получил название энтероцельного, поскольку мезодерма происходит из энтодермы при одновременном образовании целома.
У пресмыкающихся, птиц, млекопитающих животных и человека третий зародышевый листок образуется из эктодермы во время второй фазы гаструляции. Во время второй фазы гаструляции наблюдается впячивание клеток эктодермы в бластоцель при одновременной иммиграции клеток эктодермы в пространство между эктодермой и энтодермой. Клетки, выселяющиеся из эктодермы и размещающиеся между последней и слоем клеток энтодермы, являются клетками мезодермы. Этот способ образования мезодермы называется эктодермальным.
У хрящекостных рыб и земноводных наблюдается смешанный, или переходный, способ образования мезодермы, который является отражением эволюционных преобразований эмбрионального развития от низших хордовых к высшим. У этих животных мезодерма формируется в процессе гаструляции одновременно с эктодермой и энтодермой.
4. ГИСТОГЕНЕЗ И ОРГАНОГЕНЕЗ
Процесс гаструляции завершается образованием 3 зародышевых листков: наружный – эктодерма; внутренний – энтодерма и средний – мезодерма. В дальнейшем у зародыша происходит формирование нервной трубки, закладка осевых органов (хорда и сомиты), и обособление других эмбриональных зачатков.
Эмбриональные зачатки – источники развития тканей и органов в онтогенезе, представленные группами более или менее многочисленных малодифференцированных (неспециализированных) клеток; межклеточного вещества зачатки не имеют.
К эмбриональным зачаткам относятся:
1) Спинная струна, или хорда (chorda – хорда);
2) Сегментированная мезодерма, или сомиты;
3) Сегментированные ножки, или нефротом (nephros – почка);
4) Несегментированная мезодерма (боковые пластинки) или спланхнотом.
Рассмотрим, как происходит формирование нервной трубки, закладка осевых органов и обособление эмбриональных зачатков на примере птиц (рис. 3).
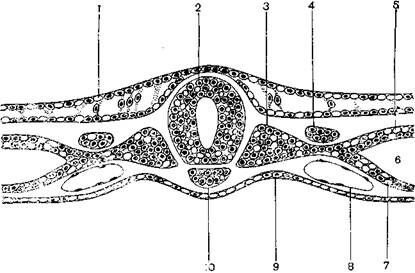
Рис. 3. Закладка осевых органов и эмбриональных зачатков у птиц (ориг.):
1 - эктодерма- 2 - нервная трубка; 3 – сомит; 4 – нефротом; 5 – париетальный листок спланхнотома; 6 – целом; 7 – висцеральный листок спланхнотома; 8 – кровяной островок; 9 – энтодерма; 10 – хорда.
Нервная трубка образуется из клеток эктодермы. Процесс образования нервной трубки у зародышей хордовых животных и человека называется нейруляцией. После образования скопления клеток мезодермы, соответствующего будущей хорде, клеточный материал эктодермы, расположенный над хордой, образует медуллярную (medulla – мозг), или нервную, пластинку. Этот обширный пласт клеток эктодермы, покрывающий хорду и сомиты, слегка прогибается, а по краям его образуются два нервных, или медуллярных, валика, представляющие собой края нервной пластинки. Прогибаясь, нервная пластинка образует нервный желобок, а нервные валики своими гребнями смыкаются, формируя нервную трубку. У бесчерепных нервная трубка на всем протяжении одинакова, она превращается в спинной мозг. У позвоночных животных и человека передний отдел нервной трубки образуется позднее за счет смыкания более высоких нервных валиков, вследствие чего головной отдел нервной трубки оказывается расширенным и из него впоследствии развиваются мозговые пузыри, а затем головной мозг,
Хорда располагается между энтодермой и нервной трубкой и имеет вид нерасчлененного сплошного клеточного тяжа. У большинства хордовых животных и человека она существует лишь у зародышей. На протяжении всей жизни хорда сохраняется у ланцетника, и некоторых рыб – цельноголовых, осетровых и двоякодышащих.
Скопление клеток мезодермы, располагающихся симметрично по бокам от хорды, представляет собой сегментированную мезодерму, или сомиты. Этому эмбриональному зачатку свойственна метамерия, т. е. расчленение спинной части мезодермы на одинаковые части, или сегменты, которые располагаются один за другим вдоль главной оси тела. Такое строение сегментированной мезодермы является характерным признаком, присущим всем хордовым.
Несегментированная мезодерма, или спланхнотом, расщепляется на два слоя клеток, или два листка, – наружный, или пристеночный, или париетальный, который примыкает к эктодерме, и внутренний, или висцеральный, который примыкает к энтодерме. Пространство, образующееся между наружным и внутренним листками спланхнотома, представляет собой вторичную полость тела. В течение некоторого времени сомиты связаны со спланхнотомом сегментными ножками, или нефротомами, но скоро связь между сегментированной и несегментированной мезодермой прерывается.
Формирование тканевых зачатков идет на основе процессов детерминации и коммитирования.
Детерминация – генетически запрограммированный путь развития клеток и тканей. В основе ее лежат стойкие изменения репрессии (блокирование) и дерепрессии (деблокирование) генов, определяющих специфику синтеза и-РНК и белков.
Коммитирование – ограничение возможных путей развития клеток.
Одновременно в первичных зачатках зародышевых и внезародышевых органов продолжаются процессы дифференцировки.
Дифференцировка – это изменения в структуре клеток, связанные с их функциональной специализацией, обусловленные активностью определенных генов.
В ходе эмбрионального развития выделяют следующие этапы дифференцировки:
I. Оотипическая дифференцировка – материал будущих зачатков представлен отдельными участками цитоплазмы яйцеклетки или зиготы.
II. Бластомерная дифференцировка – различие в клеточном материале устанавливается в бластомерах.
III. Зачатковая дифференцировка – стадия ранней гаструляции.
IV. Гистогеническая дифференцировка зачатков тканей, когда в пределах одного зародышевого листка появляются зачатки разных тканей.
Эмбриональный гистогенез – процесс возникновения специализированных тканей из малодифференцированного клеточного материала эмбриональных зачатков, происходящий в течение эмбрионального развития органов.
Гистогенез сопровождается размножением и ростом клеток, их перемещением – миграцией, дифференцировкой клеток и их производных, межклеточными и межтканевыми взаимодействиями – корреляциями, отмиранием клеток.
В процессе гистогенетической дифференцировки происходит специализация тканевых зачатков и формирование различных видов тканей. При дифференцировке клеток из исходной стволовой клетки образуются диффероны – последовательные ряды клеток. Количество дифферонов в каждом виде тканей различно.
Результатом гистогенетических процессов является формирование основных групп тканей. Их формирование начинается в эмбриональном периоде и заканчивается после рождения.
В процессе, образования многослойного зародыша последовательно формируются три полости: бластоцель, гастроцель и целом. После образования гастроцеля и целома в процессе гаструляции и нейруляции бластоцель либо сливается с гастроцелем, как, например, у земноводных, либо настолько уменьшается, что приобретает вид узких щелей, которые преобразуются в полости кровеносной системы. Гастроцель в процессе развития превращается в полость средней кишки организма, а целом образует вторичную полость тела.
5. ТЕОРИЯ ЗАРОДЫШЕВЫХ ЛИСТКОВ
Первым, кто обратил внимание на возникновение органов из зародышевых листков, или пластов, был К. Ф. Вольф (1759). Изучая развитие цыпленка, он показал, что из «неорганизованной, бесструктурной» массы яйца возникают зародышевые листки, дающие затем начало отдельным органам. К. Ф. Вольф различал нервный и кишечный листки, из которых развиваются соответствующие органы. Впоследствии X. Пандер (1817), последователь К. Ф. Вольфа, также описал наличие у куриного эмбриона зародышевых листков. К. М. Бэр (1828) обнаружил наличие зародышевых листков и у других животных, в связи с чем распространил понятие о зародышевых листках на всех позвоночных. Так, К. М. Бэр различал первичные зародышевые листки, называя их анимальным и вегетативным, из которых впоследствии, в процессе эмбрионального развития, возникают вторичные зародышевые листки, дающие начало определенным органам.
Описание зародышевых листков значительно облегчило изучение особенностей эмбрионального развития организмов и дало возможность установить филогенетические связи между животными, казалось весьма отдаленными в систематическом отношении. Это было блестяще продемонстрировано А. О. Ковалевским (1865, 1871), который по праву считается основателем современной теории зародышевых листков. А. О. Ковалевский на основании широких сравнительно-эмбриологических сопоставлений показал, что двуслойную стадию развития проходят почти все многоклеточные организмы. Он доказал сходство зародышевых листков у различных животных не только по происхождению, но и по производным зародышевых листков. Под влиянием учения о зародышевых листках Э. Геккель (1866, 1871) создал теорию гастреи, согласно которой все многоклеточные животные происходят от одного общего предка, гипотетического организма гастреи — двуслойного животного, напоминающего зародышей некоторых современных животных на стадии гаструлы либо современных кишечнополостных. Таким образом, зародышевые листки рассматривались как производные исходной двуслойной организации предков современных многоклеточных животных. В этом и состоит суть теории зародышевых листков.
Однако в теории зародышевых листков есть ряд исключении. Согласно этой теории хорда развивается из энтодермы, нервная система – из эктодермы, а мышечная ткань – из мезодермы. Однако у пресмыкающихся, птиц, млекопитающих хорда развивается из мезодермы, возникающей из эктодермы. У асцидий определенные группы бластомеров дают одновременно и хорду, и нервную систему, т. е. органы, происходящие по теории зародышевых листков из различных зародышевых листков. Гладкая мышечная ткань радужной оболочки глаза, мышц волосяных сумок кожи млекопитающих развивается не из мезодермы, как того требует теория зародышевых листков, а из эктодермы. Подобные примеры можно было бы продолжить.
Исключения из теории зародышевых листков объяснимы с точки зрения теории филэмбриогенеза А. Н. Северцова (1939). Их следует рассматривать как результат изменения ранних стадий развития, при которых не только зародышевые листки, но и сам онтогенез способны измениться до неузнаваемости в ходе эволюции под влиянием воздействий внешней среды.
Таким образом, теория зародышевых листков является крупнейшим морфологическим обобщением за всю историю эмбриологии. Благодаря ей возникло новое направление в эмбриологии, а именно эволюционная эмбриология, которая показала, что зародышевые листки, имеющиеся у подавляющего большинства животных, являются одним из свидетельств общности происхождения и единства всего животного мира.
6. ПРОИЗВОДНЫЕ ЗАРОДЫШЕВЫХ ЛИСТКОВ
С момента возникновения зародышевых листков их клеточный материал специализируется в направлении образования определенных эмбриональных зачатков, а также широкого круга тканей и органов. Уже на стадии образования зародышевых листков наблюдаются различия в их клеточном составе. Так, клетки эктодермы всегда меньше по размерам, более правильной формы и делятся быстрее, нежели клетки энтодермы. Возникающие в процессе эмбрионального развития в первично однородном материале, а также между клетками зародышевых листков различия называют дифференцировкой. Это заключительный этап эмбриогенеза.
Наружный зародышевый листок, или эктодерма, в процессе развития дает такие эмбриональные зачатки, как нервную трубку, ганглиозную пластинку, плакоды (локальные утолщения эктодермы), эктодерму кожи и внезародышевую эктодерму. Из этих эмбриональных зачатков возникают следующие ткани и органы. Нервная трубка дает нейроны и макроглию головного и спинного мозга, хвостовую мускулатуру зародышей амфибий, а также сетчатку глаза. Из ганглиозной пластинки возникают нейроны и макроглия ганглиев соматической и вегетативной нервной системы, макроглия нервов и нервных окончаний, хроматофоры низших позвоночных, птиц и млекопитающих, хромаффинные клетки, мозговой слой надпочечников, скелетные закладки челюстной, подъязычной, жаберных дуг, хрящей гортани, а также эктомезенхима. Из плакод развиваются нейроны и макроглия некоторых ганглиев, или нервных узлов, головы, а также органы равновесия, слуха и хрусталик глаза. Кожная эктодерма дает начало эпидермису кожи и его производным – железам кожи, волосяному покрову, ногтям и пр., эпителию слизистой оболочки преддверия ротовой полости, влагалища, прямой кишки и их железам, а также зубной эмали. Кроме того, из кожной эктодермы развиваются мышечные волокна волосяных сумок кожи и радужная оболочка глаза. Из внезародышевой эктодермы возникает эпителий амниона, хориона и пупочного канатика, а у зародышей пресмыкающихся и птиц – Эпителий серозной оболочки.
Внутренний зародышевый листок, или энтодерма, в развития образует такие эмбриональные зачатки, как кишечную и желточную энтодерму. Из этих эмбриональных зачатков развиваются следующие ткани и органы. Кишечная энтодерма является исходной для образования эпителия желудочно-кишечного тракта и желез – железистой части печени, поджелудочной железы, слюнных желез, а также эпителия органов дыхания и их желез. Желточная энтодерма дифференцируется в эпителий желточного мешка. Внезародышевая энтодерма развивается в соответствующую оболочку желточного мешка.
Средний зародышевый листок, или мезодерма, в процессе развития дает такие эмбриональные зачатки, как хордальный зачаток, сомиты и их производные в виде дерматома, миотома и склеротома (scleros – твердый), а также эмбриональную соединительную ткань, или мезенхиму. Кроме того, мезодерма образует нефротом, мезонефрические, или вольфовы, каналы; мюллеровы, или парамезонефрические, каналы; спланхнотом; мезенхиму, выселяющуюся из спланхнотома; внезародышевую мезодерму. Из хордального зачатка у аппендикулярий, бесчерепных, круглоротых, цельноголовых, осетровых и двоякодышащих развивается хорда, которая у перечисленных групп животных сохраняется на всю жизнь, а у позвоночных заменяется скелетогенными тканями. Дерматом дает соединительнотканную основу кожи, миотом – поперечнополосатую мышечную ткань скелетного типа, а склеротом образует скелетные ткани – хрящевую и костную. Нефротомы дают начало эпителию почки, мочевыводящих путей, а вольфовы каналы – эпителию семявыносящих путей. Мюллеровы каналы формируют эпителий яйцевода, матки и первичный эпителиальный покров влагалища. Из спланхнотома развивается целомический эпителий, или мезотелий, корковый слой надпочечников, мышечная ткань сердца и фолликулярный эпителий половых желез. Мезенхема, которая выселяется из спланхнотома, дифференцируется в клетки крови, соединительную ткань, сосуды, гладкую мышечную ткань полых внутренних органов и сосудов. Внезародышевая мезодерма дает начало соединительнотканной основе хориона, амниона, желточного мешка, а также экзоцеломическому эпителию.
Образование некоторых эмбриональных зачатков происходит раньше формирования зародышевых листков. К ним относятся трофобласт и гонобласт. Трофобласт у плацентарных млекопитающих животных и человека образуется еще при дроблении. Половой зачаток, или гонобласт, также возникает до образования трех зародышевых листков и поэтому не может быть производным какого-либо из них.
Контрольные вопросы.
1. Назовите и охарактеризуйте основные этапы эмбрионального развития, присущие всем видам животных.
2. Что называется дроблением, его типы. Чем определяется характер дробления?
3. Что называется бластулой? Какие части выделяют в бластуле зародыша? Назовите и охарактеризуйте виды бластул.
4. Какие факторы внешней среды влияют на процессы дробления?
5.Что представляет собой гаструляция и каковы особенности этого этапа эмбрионального развития у животных и человека?
6. Какие этапы выделяют в ходе гаструляции, что образуется на каждом этапе? Что образуется в процессе гаструляции?
7. Перечислите основные типы образования двухслойного и трехслойного зародыша.
8. Что послужило причиной деления животных на первичноротых и вторичноротых?
9. Что называется нейруляцией?
10. Назовите осевые органы, образующиеся в процессе нейруляции.
Люди также интересуются этой лекцией: Машина вывода и формальные системы.
11. Перечислите эмбриональные зачатки, образующиеся в эмбриогенезе.
12. Назовите основные признаки, присущие всем хордовым.
13. Какие зародышевые листки образуются в процессе эмбриогенеза?
14. Кто является автором теории зародышевых листков?
15. Что называется детерминацией, коммитированием и дифференцировкой, охарактеризуйте их.
16. Перечислите основные этапы дифференцировки, выделяющиеся в ходе эмбрионального развития.


















