
Биологические мембраны
Биологические мембраны
Строение, свойства, функции
Резюме
Биологические мембраны, наряду с цитоскелетом, формируют структуру живой клетки. Клеточная или цитоплазматическая мембрана окружает каждую клетку. Ядро окружено двумя ядерными мембранами: наружной и внутренней.
Все внутриклеточные структуры: митохондрии, эндоплазматический ретикулум, аппарат Гольджи, лизосомы, пероксисомы, фагосомы, синаптосомы и т представляют собой замкнутые мембранные везикулы (пузырьки).
Каждый тип мембран содержит специфический набор белков - рецепторов и ферментов; но основа любой мембраны - бимолекулярный слой липидов (липидный бислой), который во всякой мембране выполняет две главные функции: барьера для ионов и молекул и структурной основы ( матрицы) для функционирования рецепторов и ферментов.
Введение
Если рассмотреть электронную микрофотографию ультратонкого среза живой ткани (после его фиксации и соответствующего прокрашивания), то первое, что обращает на себя внимание, - это тонкие двойные линии, которые "вырисовывают" контуры клетки и внутриклеточных органелл.
Это - срезы через биологические мембраны - тончайшие плёнки, состоящие из двойного слоя молекул липидов и встроенных в этот слой белков. По сути дела, именно мембраны (наряду с цитоскелетом), формируют структуру живой клетки.
Клеточная или цитоплазматическая мембрана окружает каждую клетку. Ядро окружено двумя ядерными мембранами: наружной и внутренней. Все внутриклеточные структуры: митохондрии, эндоплазматический ретикулум, аппарат Гольджи, лизосомы, пероксисомы, фагосомы, синаптосомы и т представляют собой замкнутые мембранные везикулы (пузырьки).
История изучения свойств и строения мембран
Термин "мембраны" как окружающей клетку невидимой плёнки, служащей барьером между содержимым клетки и внешней средой и одновременно - полупроницаемой перегородкой, через которую могут проходить вода и некоторые растворенные в ней вещества, был впервые использован, по-видимому, ботаниками фон Молем и независимо К. фон Негели (1817-1891) в 1855 г для объясненеия явлений плазмолиза.
В 1877 г. ботаник В. Пфеффер (1845-1920) опубликовал свой труд “Исследования осмоса” (Leipzig), где постулировал существование клеточных мембран, основываясь на сходстве между клетками и осмометрами, имеющими искусственные полупроницаемые мембраны, которые были приготовлены незадолго до этого М. Траубе.
Рекомендуемые материалы
Дальнейшее изучение осмотических явлений в растительных клетках датским ботаником Х. де Фризом (1848-1935) послужило фундаментом при создании физико-химических теорий осмотического давления и электролитической диссоциации датчанином Я. Вант-Гоффом (1852-1911) и шедским ученым С. Аррениусом (1859-1922 ).
В 1888 году немецкий физико-химик В. Нернст (1864-1941) вывел уравнение диффузионного потенциала. В 1890 году немецкий физико-химик и философ В. Оствальд (1853-1932) обратил внимание на возможную роль мембран в биоэлектрических процессах.
Между 1895 и 1902 годами Э. Овертон (1865-1933) измерил проницаемость клеточной мембраны для большого числа соединений и показал прямую зависимость между способностью этих соединений проникать через мембраны и их растворимостью в липидах.
Это было чётким указанием на то,что именно липиды формируют плёнку, через которую проходят в клетку вещества из окружающего раствора.
В 1902 году Ю. Бернштейн (1839-1917) привлек для объяснения электрических свойств живых клеток мембранную гипотезу.
В 1925 году Гортер и Грендел показали, что площадь монослоя липидов, экстрагированных из мембран эритроцитов, в два раза больше суммарной площади эритроцитов. Гортер и Грендел экстрагировали липиды из гемолизированных эритроцитов ацетоном, затем выпаривали раствор на поверхности воды и измеряли площадь образовавшейся мономолекулярной пленки липидов.
На основе результатов этих исследований было сделано предположение, что липиды в мембране располагаются в виде бимолекулярного слоя. Это предположение подтвердили исследования электрических параметров биологических мембран (Коул и Кёртис, 1935 год): высокое электрическое сопротивление, порядка 107 Омм2 и большая электроемкость 0,51 '/м2.
Вместе с тем имелись экспериментальные данные, которые свидетельствовали о том, что биологическая мембрана содержит в своем составе и белковые молекулы.
Эти противоречия экспериментальных результатов были устранены Даниелли и Давсоном, предложившими в 1935 году так сказать "бутербродную" модель строения биологических мембран, которая с некоторыми несущественными изменениями продержалась в мембранологии в течении почти 40 лет. Согласно этой модели, на поверхности фосфолипидного бислоя в мембранах располагаются белки.
Функции биологических мембран
В таблице 1 перечислены функции цитоплазматических и некоторых внутриклеточных мембран.
Во всех живых клетках биологические мембрану выполняют функцию барьера, отделяющего клетку от окружающей среды, и разделяющего внутренний объем клетки на сравнительно изолированные "отсеки" (compartments).
Сами по себе перегородки, разделяющие клетки на отсеки, построены из двойного слоя липидных молекул (называемого часто липидным бислоем) и практически непроницаемы для ионов и полярных молекул, растворимых в воде.
Но в этот липидный бислой встроены многочисленные белковые молекулы и молекулярные комплексы, одни из которых обладают свойствами селективных (т. е. избирательных) каналов для ионов и молекул, а другие - насосов, способных активно перекачивать ионы через мембрану. Барьерные свойства мембран и работа мембранных насосов создают неравновестное распределение ионов между клеткой и внеклеточной средой, что лежит в основе процессов внутриклеточной регуляции и передачи сигналов в форме электрического импульса между клетками.
Вторая функция, общая для всех мембран - это функция "монтажной платы" или матрицы, на которой располагаются в определенном порядке белки и белковые ансамбли, образующие системы переноса электронов, запасания энергии в форме АТФ, регуляции внутриклеточных процессов гормонами, поступающими извне и внутриклеточными медиаторами, узнавания других клеток и чужеродных белков, рецепции света и механических воздействий и т
Гибкая и эластичная пленка, которой по существу являются все мембраны, выполняет и определенную механическую функцию, сохраняя клетку целой при умеренных механических нагрузках и нарушениях осмотического равновесия между клеткой и окружающей средой.
Общие для всех мембран функции барьера для ионов и молекул и матрицы для белковых ансамблей обеспечиваются главным образом липидным бислоем, который устроен в принципе одинаково во всех мембранах.
Однако набор белков индивидуален для каждого типа мембран, что позволяет мембранам участвовать в выполнении самых разных функций в различных клетках и клеточных структурах. Некоторые из этих фукнкций упомянуты в таблице 1.
Строение мембран
Общая схема строения мембран
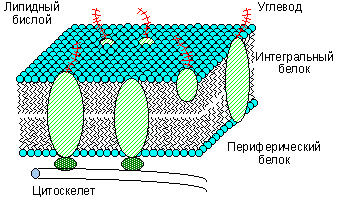
Согласно современным предтавлениям, все клеточные и внутриклеточные мембраны устроены сходным образом: основу мембраны составляет двойной молекулярный слой липидов (липидный бислой) на котором и в толще которого находятся белки ( см. рис. 1).
Липиды мембран
Липидные бислои образуются амфифильными молекулами фосфолипидов и сфингомиелина в водной фазе. Амфифильными эти молекулы называют потому, что они состоят из двух частей, различных по своей растворимости в воде: полярной "головки", обладающей высоким сродством к воде, т. е. гидрофильной, и "хвоста", образуемого неполярными углеводородными цепями жирных кислот; эта часть молекулы обладает низким сродством к воде, т. е. гидрофобна.
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин. Например, в мембранах эритроцитов человека их содержание, составляет, соответственно 36, 30 и 22 % по весу; еще 12% приходится на гликолипиды (Котык А. и Яначек К. Мембранный транспорт, Москва, МИР, 1980 г., стр. 45).
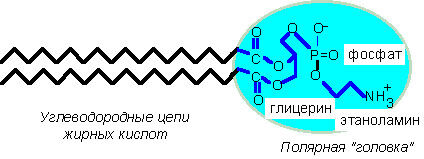
Примером амфифильной молекулы может служить молекула фосфатидилэтаноламина, структура которой показана на рис. 2. Как и другие фосфолипиды, фосфатидилэтаноламин, в химическом отношении представляет собой сложные эфиры трехатомного спирта глицерина с двумя жирными кислотами; к третьей гидроксильной группе присоединен ортофосфат, а к нему - небольшая органическая молекула, характерная для каждого вида фосфолипидов. В рассматриваемом случае это этаноламин, но могут быть также холин, инозитол, серин и некоторые другие молекулы.
В состав липидного слоя мембран входят также холестерин и сфингомиелины; последние близки к фосфолипидам по химическому строению и физическим свойствами.
Химия фосфолипидов
С химической точки зрения фосфолипид состоит из четырёх частей: глицерина, двух жирных кислот с длинной углеводородной цепью, фосфорной кислоты и особой для каждого фосфолипида группы, которую мы будем называть характеристической группой. Трёхатомный спирт - глицерин связывает через сложно-эфирную связь две жирные кислоты и остаток фосфорной кислоты
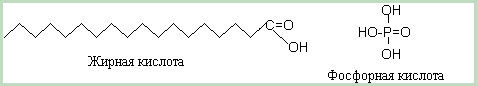
Рис. 4. Скелетные формулы одной из жирных кислот и фосфорной кислоты
Фосфолипиды различаются как составом жирных кислот, так и структурой характеристической группы. В фосфатидилэтаноламине такой группой является остаток этаноламина. В других фосфолипидах такой группой может быть остаток холина, серина и другие полярные молекулы (см. рис. 5).

Рис. 5. Характеристические (полярные) группы фосфолипидов
Белки мембран
Белки мембран принято делить на интегральные и периферические. Интегральные белки имеют обширные гидрофобные участки на поверхности и нераствориммы в воде.
С липидами мембран они связаны гидрофобными взаимодействиями и частично погружены в толщу липидного бислоя, а зачастую и пронизывают бислой, оставляя на поверхности сранительно небольшие гидрофильные участки.
Отделить эти белки от мембраны удается только с помощью детергентов, типа додецилсульфата или солей желчных кислот, которые разрушают липидный слой и переводят белок в растворимую форму (солюбилизируют его) образуя с ним ассоциаты. Все дальнейшие операции по очистке интегральных белков осуществляются также в присутствии детергентов.
Периферические белки связаны с поверхностью липидного бислоя электростатическими силами и могут быть отмыты от мембраны солевыми растворами.
Самосборка мембранных структур
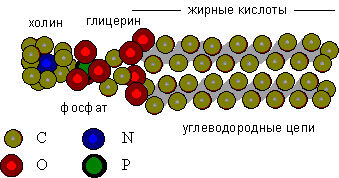
Данные рентгеноструктурного анализа и другие показавают, что молекулы фосфолипидов имеют форму сплюснутого с боков цилиндра, а по длине как бы делятся на две неравные части: небольшую "голову", состоящую из полярных групп, и длинный "хвост", образованный углеводородными цепями жирных кислот, входящих в состав фосфолипида (см. Рис. 6).
|
| Рис. 6. Молекулярная структура фосфатидилхолина (лецитина) Левая часть молекулы, содержащая атомы кислорода, фосфора и азота и заряженные группы (триметиламин и фосфат) активно взаимодействует с молекулами воды (гидрофильны); правая часть состоит из углеводородных цепей жирных кислот, и отталкивает воду (гидрофобна). |
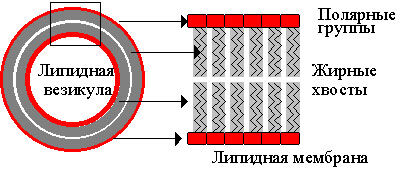
Такое строение молекулы приводит к тому, что в водных растворах фосфолипидные молекулы самособираются в бислойную мембрану. В мембране "жирные хвосты" упрятаны внутрь, а снаружи в контакте с водным окружением оказываются полярные "головы" этих молекул.
|
| Рис. 7. Самосборка фосфолипидных молекул в липидных везикулы в водном растворе. Каждая фосфолипидная молекула состоит из полярной группы и жирнокислотных хвостов. В водном растворе происходит самосборка мембран (справа) и замыкание мембран с образованием липидных пузырьков, называемых липосомами (слева). |
Модельные мембраны. Липосомы
Изучение физических свойств липидного слоя мембран осуществляется преимущественно на двух видах искусственных мембранных структур, образованных синтетическими фосфолипидами или липидами, выделенными из биологических источников: липосомах и бислойных липидных мембранах (БЛМ).
Липосомы - это липидные везикулы (пузырьки), образующиеся из фосфолипидов в водных растворах. Чтобы получить липосомы, спиртовый раствор фосфолипидов впрыскивают в большой объем водного раствора.Фосфолипиды, нерастворимые в воде, образуют мелкие пузырьки, стенки которых состоят из одного липидного бислоя (однослойные липосомы).
|
| Рис. 7. Самосборка фосфолипидных везикул в водном растворе Каждая фосфолипидная молекула состоит из полярной головки и жирнокислотных хвостов. В водном растворе головки выставляются науржу, а хвосты прячутся внутрь; получаются мембранные везикулы. |
Можно сначала высушить раствор фосфолипидов в органическом растворителе (например, хлороформе) в пробирке, добавить в пробирку водный раствор и хорошенько потрясти пробирку. Липиды переходят в водный раствор, теперь уже в виде многослойных липосом.
Суспензию липосом обычно используют для изучения физических свойств липидного бислоя как вязкость, поверхностный заряд или диэлектрическая проницаемость, а также для изучения проницаемости для незаряженных молекул.
БЛМ
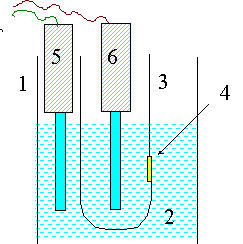
Для изучения ионной проницаемости липидного слоя мембран используют БЛМ. Для приготовления БЛМ (см. рис. 8) в стаканчик с раствором электролита помещают второй, тефлоновый стаканчик , в стенку которого сделано отверстие, диаметром около 1 мм.
|
| Бесплатная лекция: "9. Кинетика ферментативных реакций" также доступна. Рис. 8. Приготовление бимолекулярных липидных мембран (БЛМ) В стеклянный стакан (1) помещают раствор электролита (2) и опускают тефлоновый сосуд 3 с отверстием в стенке (4). В отверстии формируют БЛМ (см. следующий рисунок). |
В такой пленке полярные головки фосфолипидов обращены в водную фазу, а неполярные углеводородные цепи жирных кислот сливаются в сплошную вязкую фазу во внутренней части липидной мембраны. По многим свойствам эта пленка сходна с липидным слоем биологических мембран.
|
| Рис. 9. Образование БЛМ в отверстии в стенке тефлонового сосуда A - вносим с помощью капилляра (4) каплю раствора фосфолипида в гептане (5) в отверстие в стенке сосуда (3). |
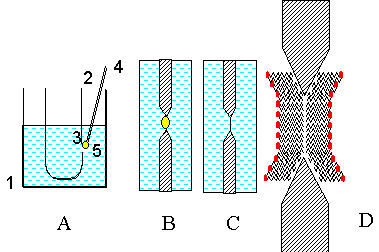
Если липосомы широко используются для изучения таких свойств липидного слоя мембран как микровязкость (методом флуоресцентных и спиновых зондов), фазовые переходы в липидах (методом микрокалориметрии) и химические реакции в липидном слое, то БЛМ идеально подходит для изучения ионной проницаемости путем измерения электрической проводимости мембраны и образующихся на ней потенциалов.


















