


Часть вторая
2. Система обратных связей
(Обратные связи обычно носят отрицательный характер):
• Усиление выработки гормонов периферическими железами угнетает, а ослабление – стимулирует секрецию соответствующих факторов гипоталамуса и тропных гормонов гипофиза.
• В клетках-мишенях нарабатываются метаболиты-индикаторы, которые также по механизму обратной связи могут блокировать свой синтез на уровне эндокринных желез и гипоталамуса.
(Рис) Система обратных связей на примере ГГНС (гипоталамус-гипофиз-кора надпочечников)
Нейроны гипоталамуса синтезируют кортиколиберин, он попадает в переднюю долю гипофиза. Нейроны гипофиза в ответ выделяют кортикотропин (АКТГ), он стимулирует секрецию кортикостероидов в коре надпочечников. Уровень кортикостероидов в крови действует как сигнал обратной связи и заставляет гипофиз или ЦНС в целом продолжать, или приостанавливать весь этот процесс синтеза.
 Последовательность событий:
Последовательность событий:
1) Поток информации о состоянии внешней и внутренней среды организма (анализаторы, термо-и хеморецепторы, внутренняя среда) поступает в ЦНС, преобразуется в нервные импульсы и передается через синапсы с помощью химических сигналов-медиаторов на эффекторные клетки, изменяя их метаболизм. Гипоталамус – часть ЦНС и связан с другими структурами ЦНС (спинной, средний, продолговатый мозг, таламус, базальные ядра, лимбические образования, ряд полей коры больших полушарий и др.).
2) Одновременно, гипоталамус – центральный орган эндокринной системы, он объединяет 2 потока информации (нервный и гуморальный). Нейросекреторные клетки гипоталамуса преобразуют афферентные нервные стимулы в гормональные факторы – нейропептиды (9 либеринов (+), 4 статина (-)), которые регулируют →
Рекомендуемые материалы
3) выделение передней долей гипофиза тропных гормонов, которые влияют на →
4) секрецию в периферических эндокринных железах гормонов, меняющих →
5) метаболизм в клетках-мишенях
Эффект эндокринной регуляции реализуется через изменения:
· активности уже существующих в клетке белков (ферментов, рецепторов, факторов транскрипции, трансляции и т.д.) путем ковалентной или аллостерической модификации, взаимодействия субъединиц и др.
· количества белков (индукция/репрессия синтеза или разрушение: чаще всего – транскрипция и процессинг иРНК, трансляция и процессинг белков)
· скорости транспорта веществ через мембраны (перемещение внутриклеточных сигналов: например, меняется компартментализация белков-эффекторов)
ИТАК: Эндокринные железы и их гормоны тесно взаимодействуют с нервной системой, образуя общий механизм регуляции.
Определяющая роль у ЦНС, а гормоны – интегрирующие регуляторы, связывают различные регуляторные механизмы и метаболизм в разных органах. Они функционируют как первичные химические посредники, передающие сигналы, возникающие в органах и ЦНС.
ВАЖНО: Все уровни регуляции тесно взаимосвязаны, конечной точкой любого пути регуляции является изменение активности белка (фермента).
Термин "гормон" ввёл Старлинг в 1905 г. (от лат hormanio – побуждаю)
для обозначения химических соединений, вырабатываемых в эндокринных железах, выделяемых в кровь или лимфу (10-9–10-12 М), обладающих высокой биологической активностью, регулирующих обмен веществ и развитие организма.
1 молекула адреналина способствует высвобождению из гликогена 108 молекул глюкозы
Классификации гормонов
I. по химическому строению:
1. производные аминокислот (из ФЕН, ТИР – адреналин, норадреналин, тиреоиды)
2. белково-пептидные (инсулин, малые регуляторные пептиды, ПТГ, кальцитонин)
3. стероидные (половые, глюко-, минералокортикоиды, из вит Д3 – кальцитриол)
4. производные высших жирных кислот (простагландины, тромбоксаны, простациклины и лейкотриены) синтез по цикло- и липо- оксигеназным механизмам
На примере надпочечников – фактически это 2 железы внутренней секреции:
1) внутренний слой – мозговая часть, формируется из симпатического ганглия (адреналовой ткани) и вырабатывает из тирозина катехоламины (биогенные амины, содержащие общий фрагмент: 3,4-дигидроксифенилаланин («катехол»)). 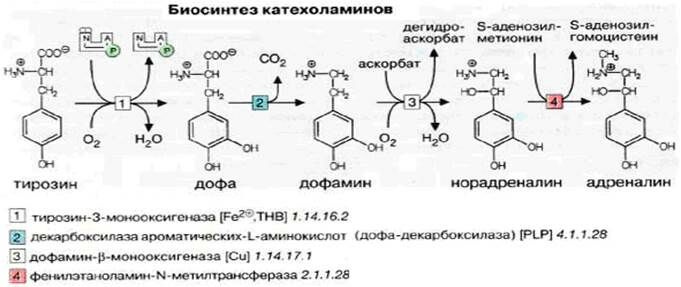
· При гидроксилировании тир [1] образуется дофа (3,4-дигидроксифенилаланина).
· При декарбоксировании [2] дофа образуется дофамин.
· При гидроксилировании [3] дофамина образуется норадреналин (норэпинефрин).
Донор водорода в этой реакции аскорбат.
· Метилирование [4] норадреналина ведёт к образованию адреналина (эпинефрин).
Донор метильной группы в этой реакции SАМ.
2) наружный слой – корковый, трехслойный, образуется из мезодермы, из которой также закладываются гонады. Этим объясняется химическое сродство гормонов коры надпочечников и половых гормонов – стероиды. На долю коры приходится ~ 80% массы железы, из неё выделено ~ 50 стероидов.
Биосинтез кортикостероидов из холестерола в коре надпочечников
++++++++++++++++++++++++666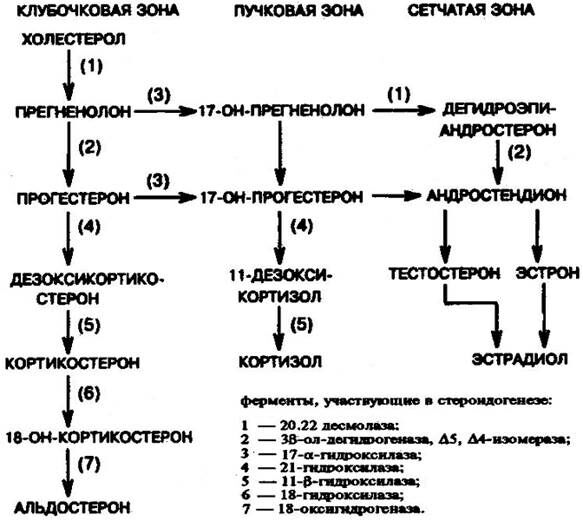
!!!!!! Знать основные принципы синтеза стероидов из холестерола и формулы: прогестерона, эстрадиола, тестостерона, кортизола и альдостерона (в учебнике).
II. по функции:
1. регулирующие синтез и секрецию гормонов эндокринных желез (либерины и статины гипоталамуса, тропные гормоны гипофиза, пролактин, ФСГ, ЛГ и др.).
2. обмена кальция и фосфатов (ПТГ, кальцитонин, кальцитриол)
3. водно-солевого обмена (альдостерон, АДГ)
4. репродуктивной системы (андрогены, гестагены, эстрогены)
5. обмена углеводов, липидов, АК (инсулин, глюкагон, глюкокортикоиды)
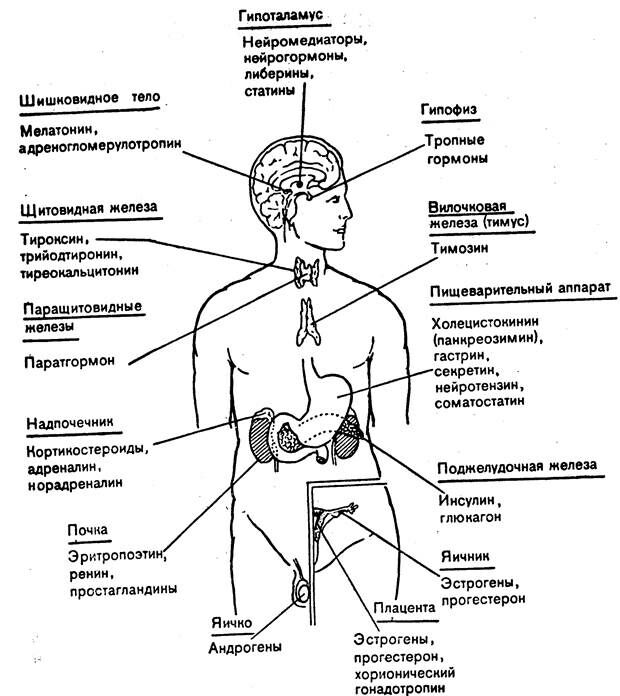
III. по принадлежности к эндокринным железам
(место образования - отдельные органы или группы клеток) – см рисунок ниже
Свойства гормонов:
1. дистантность действия (преимущественно, но могут действовать местно),
2. специфичность (рецепторы) – у каждого свой эффект (иногда – неспецифически),
3. высокая биоактивность – эффект от минимальных концентраций (10-8 – 10-12 М), секреция в небольших количествах (0,2–20,0 мг/сут) зависит от потребности организма: эстрогены 0,25 мг/сут, инсулин 1,5 мг/сут, андрогены 5,0 мг/сут, глюкокортикоиды 20,0 мг/сут, минералокортикоиды 2,0 мг/сут, тиреоиды 0,3 мг/сут; каскадное усиление конеч. эффекта
4. кратковременность действия – высокая скорость образования/инактивации (период полураспада АКТГ ~ 5 мин, инсулин, андрогены и эстрогены – до 30 мин; тироксин – до 3-4 сут)
5. дозозависимость (вплоть до противоположных эффектов при разных дозах) и возможность разнонаправленного действия в разных тканях
6. посредническая роль – в передаче информации НС→клетка (обеспечивают гуморальный, частично – контактный межклеточный контроль за метаболическими, структурными и физиологическими процессами на различных уровнях организации организма).
Этапы реализации действия гормонов (ВЫДЕЛЕНО ЖЁЛТЫМ!)
· 1) Регуляция синтеза гормонов в железах внутренней секреции
(контроль нервной системы – прямым иннервационным путём;
тропная регуляция – системой гипоталамус–гипофиз;
механизм обратной связи – отрицательным влиянием периферических гормонов
на активность верхних отделов эндокринной системы)
· 2) Синтез и «созревание» молекул гормонов
· 3) Депонирование и секреция (постоянная – тиреоиды; эпизодическая – ЛГ; периодическая – АКТГ, кортизол; в ответ на какие-либо стимулы – инсулин (изменение концентрации глюкозы), паратгормон и кальцитонин (изменение концентрации Са2+))
· 4) Транспорт в крови. Свободные и связанные с белками гормоны (особые транспортные белки (тироксин-связывающий, сексстероид-связывающий глобулины и др.))
· 5) Эффекторное звено: взаимоотношения с рецепторами
· 6) Период полужизни гормональных молекул, биотрансформация и выведение
(для пептидов и белков – протеолиз, для стероидов – микросомальное окисление и образование коньюгатов (сульфатов или глюкуронидов))
· 7) Взаимоотношения с другими регуляторами и эффекторами
(синергисты, антагонисты)
Механизмы передачи гормонального сигнала в клетку (трансдукция)
1. Мембранно-внутриклеточный (действие гидрофильных гормонов).
Передача сигнала в клетку через мембранные рецепторы (проводники сигнала). Рецепторы – интегральные белки мембран. На наружной поверхности плазмалеммы образуется гормон-рецепторный комплекс, что ведёт к изменению пространственной структуры белка. Это генерирует новый сигнал на внутренней стороне мембраны по одному из 3 механизмов, поэтому → 3 типа рецепторов
Типы рецепторов:
1) Рецепторы первого типа – трансмембранные аллостерические ферменты из одной полипептидной цепи, активный центр расположен на внутренней стороне мембраны. К этому типу принадлежат рецепторы инсулина.
Связывание сигнального вещества (гормона) ведет к димеризации рецептора. При этом фермент активируется и фосфорилирует остатки тирозина в ряде белков (GluT и др.). В первую очередь – автофосфорилирование молекулы самого рецептора, после чего с ней связывается белок-переносчик сигнала к внутриклеточным протеинкиназам.
2) Рецепторы второго типа – олигомерные мембранные белки, образующие лиганд-активируемый ионный канал. Связывание лиганда (гормона) ведет к открыванию канала для ионов Na+, К+ или Cl-. По такому механизму осуществляется действие нейромедиаторов: ацетилхолин (никотиновые рецепторы: Na+- и К+-каналы), ГАМК (А-рецептор: Cl--канал), ангиотензин и др.
3) Рецепторы третьего типа – сопряжены с ГТФ-связывающими белками. Полипептидная цепь рецептора включает семь трансмембранных тяжей. Рецепторы передают сигнал с помощью ГТФ-связывающих белков G-белков
(Gs – стимулирует аденилатциклазу, Gi – ингибирует аденилатциклазу, Gq – активирует фосфолипазу С) на белки-эффекторы (ферменты или ионные каналы), которые изменяют концентрацию вторичных посредников (мессенджеров) или ионов.
Например, Gs активирует аденилатциклазу, локализованную на внутренней стороне мембраны и катализирующую превращение АТФ в цАМФ.
Схема мембранно-внутриклеточного действия гормонов на метаболизм клетки

СХЕМА. Этапы преобразования сигнала G-белками с передачей на циклоАМФ
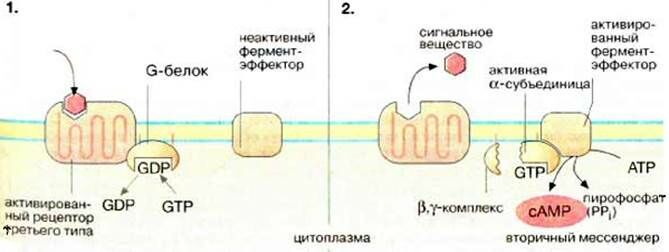
При связывании гормона с рецептором третьего типа конформация рецептора меняется и Г-Р комплекс приобретает способность присоединять G-белок. В неактивном состоянии G-белок связан с ГДФ. Ассоциация G-белка с рецептором ведёт к обмену ГДФ на ГТФ, G-белок активируется, отделяется от рецептора, диссоциирует на 2 части. Активная α-субъединица G-белка переносит сигнал на белки-эффекторы. Если белок-эффектор – аденилатциклаза, то он преобразует АТФ в цАМФ. Медленный гидролиз связанного с α-субъединицей ГТФ до ГДФ переводит активную α-субъединицу G-белка в неактивное состояние и она вновь ассоциирует с остальной βγ-частью молекулы → G-белок возвращается в исходное состояние. цАМФ нарабатывается в клетку, пока сохраняется Г-Р комплекс
(1 молекула активного комплекса способствует образованию от 10 до 100 цАМФ).
8 Использование графики, цвета и текста - лекция, которая пользуется популярностью у тех, кто читал эту лекцию.
СХЕМА. Активация 2-х форм гуанилатциклазы первичными мессенджерами

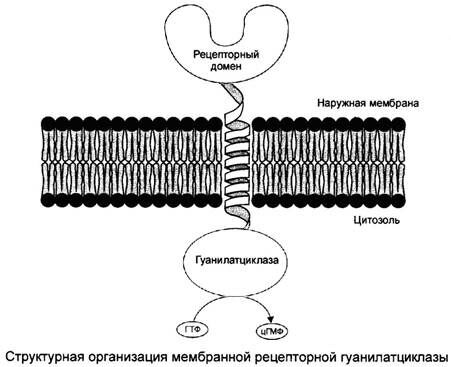
вариант рисунка для представления материала.
Преобразование сигнала G-белками: передача на циклоАМФ или цикло ГМФ.


















