Лекция. Общая характеристика функций нервной системы. Нейрон как структурно-функциональная единица ЦНС. Физиология синапсов. Механизмы передачи возбуждения через синапсы
Описание
Тема лекции: Общая характеристика функций нервной системы. Нейрон как структурно-функциональная единица ЦНС. Физиология синапсов. Механизмы передачи возбуждения через синапсы
План лекции
Центральная нервная система (ЦНС)![]() Рис.1. Строение центральной нервной системы.
Рис.1. Строение центральной нервной системы.
А -головной и спинной мозг со спинальными нервами. Б - основные компоненты центральной нервной системы. Показаны также четыре главные доли коры больших полушарий: затылочная, теменная, лобная и височная
![]() Рис. 2. Головной мозг. А - среднесагиттальный срез головного мозга. Обратите внимание на относительное расположение коры больших полушарий, мозжечка, таламуса и ствола мозга, а также различных комиссур. Б и В - система желудочков мозга in situ - вид сбоку (Б) и спереди (В)
Рис. 2. Головной мозг. А - среднесагиттальный срез головного мозга. Обратите внимание на относительное расположение коры больших полушарий, мозжечка, таламуса и ствола мозга, а также различных комиссур. Б и В - система желудочков мозга in situ - вид сбоку (Б) и спереди (В)
Тест
1. ЦНС представлена
a) спинным и головным мозгом
b) головным мозгом
2. Выберите наиболее правильный ответ. Основная роль ЦНМ заключается в
a) переработке информации от ПНС и выработке программы действия
b) регуляции движением
Функциональная единица нервной системы – нейрон (рис 3).
Типичный нейрон обладает воспринимающей поверхностью в виде тела (сомы) и нескольких отростков – дендритов, на которых находятся синапсы, т.е. межнейронные контакты. Аксон нервной клетки образует синаптические связи с другими нейронами или с эффекторными клетками. Коммуникативные сети нервной системы складываются из нейронных цепей, образованных синаптически взаимосвязанными нейронами.
![]() Рис. 3. Схема «идеального» нейрона и его основных компонентов.
Рис. 3. Схема «идеального» нейрона и его основных компонентов.
Большинство афферентных входов, поступающих по аксонам других клеток, оканчиваются синапсами на дендритах (Д), но некоторые - синапсами на соме. Возбуждающие нервные окончания чаще располагаются дистально на дендритах, а тормозные нервные окончания чаще находятся на соме.
Сома. В соме нейронов находятся ядро и ядрышко (рис. 3), а также хорошо развитый аппарат биосинтеза, который производит компоненты мембран, синтезирует ферменты и другие химические соединения, необходимые для специализированных функций нервных клеток. К аппарату биосинтеза в нейронах относятся тельца Ниссля - плотно прилегающие друг к другу сплюснутые цистерны гранулярного эндоплазматического ретикулума, а также хорошо выраженный аппарат Гольджи. Кроме того, сома содержит многочисленные митохондрии и элементы цитоскелета, в том числе нейрофиламенты и микротрубочки. В результате неполной деградации мембранных компонентов образуется пигмент липофусцин, накапливающийся с возрастом в ряде нейронов. В некоторых группах нейронов ствола мозга (например, в нейронах черной субстанции и голубого пятна) заметен пигмент мелатонин.
Дендриты, выросты клеточного тела, у некоторых нейронов достигают длины более 1 мм, и на их долю приходится более 90% площади поверхности нейрона. В проксимальных частях дендритов (ближе к клеточному телу) содержатся тельца Ниссля и участки аппарата Гольджи. Однако главные компоненты цитоплазмы дендритов - микротрубочки и нейрофиламенты. Дендриты многих нейронов обладают потенциалуправляемой проводимостью. Часто это обусловлено присутствием кальциевых каналов, при активации которых генерируются кальциевые потенциалы действия.
Аксон. Специализированный участок тела клетки (чаще сомы, но иногда - дендрита), от которого отходит аксон, называется аксонным холмиком. Аксон и аксонный холмик отличаются от сомы и проксимальных участков дендритов тем, что в них нет гранулярного эндоплазматического ретикулума, свободных рибосом и аппарата Гольджи. В аксоне присутствуют гладкий эндоплазматический ретикулум и выраженный цитоскелет.
Нейроны сообщаются друг с другом с помощью потенциалов действия, распространяющихся в нейронных цепях по аксонам. Потенциалы действия поступают от одного нейрона к следующему в результате синаптической передачи. В процессе передачи достигший пресинаптического окончания потенциал действия обычно запускает высвобождение нейромедиатора, который либо возбуждает постсинаптическую клетку, так что в ней возникает потенциал действия, либо тормозит ее активность. Аксоны не только передают информацию в нейронных цепях, но и доставляют путем аксонального транспорта химические вещества к синаптическим окончаниям.
Органеллы нейрона. На рисунке 4 представлена сома нейронов. В соме нейронов показаны ядро и ядрышко, аппарат биосинтеза, который производит компоненты мембран, синтезирует ферменты и другие химические соединения, необходимые для специализированных функций нервных клеток. В него входят тельца Ниссля - цистерны гранулярного эндоплазматического ретикулума, а также хорошо выраженный аппарат Гольджи. Сома содержит митохондрии и элементы цитоскелета, в том числе нейрофиламенты и микротрубочки. В результате неполной деградации мембранных компонентов образуется пигмент липофусцин, накапливающийся с возрастом в ряде нейронов. В некоторых группах нейронов ствола мозга (например, в нейронах черной субстанции и голубого пятна) заметен пигмент мелатонин.
![]() Рис. 4. Нейрон. А - органеллы нейрона. На поверхности нейрона - синаптические окончания. Б - Схема соответствует электронно-микроскопической картине. Видны ядро, ядрышко, хроматин, ядерные поры. В цитоплазме - митохондрии, шероховатый эндоплазматический ретикулум, аппарат Гольджи, нейрофиламенты и микротрубочки. На наружной стороне плазматической мембраны - синаптические окончания и отростки астроцитов.
Рис. 4. Нейрон. А - органеллы нейрона. На поверхности нейрона - синаптические окончания. Б - Схема соответствует электронно-микроскопической картине. Видны ядро, ядрышко, хроматин, ядерные поры. В цитоплазме - митохондрии, шероховатый эндоплазматический ретикулум, аппарат Гольджи, нейрофиламенты и микротрубочки. На наружной стороне плазматической мембраны - синаптические окончания и отростки астроцитов.
Типы нейронов
Нейроны разного типа выполняют специфичные коммуникативные функции (рис. 5).
![]() Рис. 5. Типы нейронов
Рис. 5. Типы нейронов
А - нейроны разнообразной формы: 1 - нейрон, напоминающий пирамиду (пирамидные клетки коры больших полушарий); 2 - клетки Пуркинье в коре мозжечка; 3 - постганглионарный симпатический мотонейрон; 4 - альфа-мотонейрон спинного мозга, мультиполярный, с радиальными дендритами; 5 - сенсорная клетка спинального ганглия; не имеет дендритов, ее отросток разделяется на две ветви: центральную и периферическую, псевдоуниполярные нейроны. Б - типы нейронов
Так, нейроны ганглиев задних корешков (спинальных ганглиев) получают информацию от сенсорных нервных окончаний в органах. Их клеточные тела лишены дендритов (рис. 5 ) и не получают синаптических окончаний. Выйдя из клеточного тела, аксон такого нейрона разделяется на две ветви, одна из которых (периферический отросток) направляется в составе периферического нерва к сенсорному рецептору, а другая ветвь (центральный отросток) входит в спинной мозг (в составе заднего корешка) либо в ствол мозга (в составе черепного нерва).
Нейроны другого типа, такие, как пирамидные клетки коры больших полушарий и клетки Пуркинье коры мозжечка, заняты переработкой информации (рис. 5). Их дендриты покрыты дендритными шипиками и характеризуются обширной поверхностью. Они имеют огромное количество синаптических входов.
Виды ненейронных клеток (нейроглия)
Нейроглии (рис. 6) в ЦНС человека на порядок больше, чем число нейронов: 1013 и 1012 соответственно. Нейроглия способствует осуществлению коммуникативных процессов нейронами, образует миелиновую оболочку, увеличивающую скорость проведения потенциалов действия.
Типы нейроглии
В ЦНС к нейроглии относят астроциты и олигодендроциты, клетки микроглии и эпендимы, а в ПНС - шванновские клетки и клетки-сателлиты.
![]() Рис. 6. Ненейронные клетки. А - схематическое представление ненейронных элементов ЦНС: два астроцита, ножки отростков которых заканчиваются на соме и дендритах нейрона, а также контактируют с мягкой мозговой оболочкой и/или капиллярами; олигодендроцит формирует миелиновую оболочку аксонов; клетки микроглии и клетки эпендимы. Б - разные типы клеток нейроглии в ЦНС: 1 - фибриллярный астроцит; 2 - протоплазматический астроцит; 3 - олигодендроцит; 4 - клетки микроглии; 5 - клетки эпендимы
Рис. 6. Ненейронные клетки. А - схематическое представление ненейронных элементов ЦНС: два астроцита, ножки отростков которых заканчиваются на соме и дендритах нейрона, а также контактируют с мягкой мозговой оболочкой и/или капиллярами; олигодендроцит формирует миелиновую оболочку аксонов; клетки микроглии и клетки эпендимы. Б - разные типы клеток нейроглии в ЦНС: 1 - фибриллярный астроцит; 2 - протоплазматический астроцит; 3 - олигодендроцит; 4 - клетки микроглии; 5 - клетки эпендимы
Астроциты (звездчатая форма) регулируют микросреду вокруг нейронов ЦНС, их отростками окружены группы синаптических окончаний, которые в результате изолированы от соседних синапсов. Особые отростки - «ножки» астроцитов образуют контакты с капиллярами и с соединительной тканью на поверхности ЦНС - с мягкой мозговой оболочкой (рис. 6). Ножки ограничивают свободную диффузию веществ в ЦНС. Астроциты могут активно поглощать К+и нейромедиаторные вещества, затем метаболизируя их. Таким образом, астроциты играют буферную роль, перекрывая прямой доступ для ионов и нейромедиаторов во внеклеточную среду вокруг нейронов. В цитоплазме астроцитов находятся глиальные филаменты, выполняющие в ткани ЦНС механическую опорную функцию. В случае повреждения отростки астроцитов, содержащие глиальные филаменты, подвергаются гипертрофии и формируют глиальный «рубец».
Другие элементы нейроглии обеспечивают электрическую изоляцию нейронных аксонов. Многие аксоны покрыты изолирующей миелиновой оболочкой. В ЦНС миелиновую оболочку создают мембраны клеток олигодендроглии (рис. 6). В ПНС миелиновая оболочка образована мембранами шванновских клеток (рис. 6). Немиелинизированные (безмякотные) аксоны ЦНС не имеют изолирующего покрытия.
Миелин увеличивает скорость проведения потенциалов действия благодаря тому, что ионные токи во время потенциала действия входят и выходят только в перехватах Ранвье (областях перерыва между соседними миелинизирующими клетками). Таким образом, потенциал действия «перескакивает» от перехвата к перехвату - так называемое сальтаторное проведение.
Кроме того, в состав нейроглии входят клетки-сателлиты, инкапсулирующие нейроны ганглиев спинальных и черепных нервов, регулируя микросреду вокруг этих нейронов наподобие того, как это делают астроциты.
Еще один вид клеток - микроглия, или латентные фагоциты. В случае повреждения клеток ЦНС микроглия способствует удалению продуктов клеточного распада. В этом процессе участвуют другие клетки нейроглии, а также фагоциты, проникающие в ЦНС из кровотока.
Ткань ЦНС отделена от ЦСЖ, заполняющей желудочки мозга, эпителием, сформированным эпендимными клетками (рис. 6). Эпендима обеспечивает диффузию многих веществ между внеклеточным пространством мозга и ЦСЖ. Специализированные эпендимные клетки сосудистых сплетений в системе желудочков секретируют значительную долю ЦСЖ.
Тест
1. Выберите наиболее точный ответ. В ЦНС присутствуют клетки, обеспечивающих функцию анализа информации и передачи ее эффекторным клеткам
a) нейроны и глиальные клетки
b) различные типы нейронов
2. Верно ли, что миелиновая оболочка нервных волокон обеспечивает более высокую скорость проведения возбуждения
a) да
b) нет
3. Глиальные клетки выполняют
a) разнообразные функции
b) только функцию опоры и питания для нейронных сетей
Электрическое взаимодействие клеток: электрический синапс
На рисунке 7А схематично представлена клетка с основными известными типами контактных структур.
1. Плотный контакт (tight junction) характеризуется слиянием внешних листков мембран соседних клеток с образованием одиночной электронно-плотной полосы вдоль контакта и представляет собой сеть ветвящихся тонких гребней
2. Непосредственно к плотному контакту примыкает зона слипания, или промежуточный контакт (intermediate junction), заполненный гомогенным материалом низкой электронной плотности.
3. Десмосома (desmosome) - локальное дискообразное электронно-плотное образование, располагающееся параллельно внутреннему листку каждой клеточной мембраны. От каждой десмосомы внутрь цитоплазмы расходятся пучки фибрилл.
4. Щелевой контакт (gap junction) - область тесного сближения мембран (20-4-А) двух соседних клеток. Представляет интерес для физиологии с позиций межклеточного взаимодействия.
Щелевой контакт, или gap junction, - тип контактов между клетками как электровозбудимых, так и электроневозбудимых тканей. В бислое мембран обеих клеток, образующих щелевой контакт, белковая часть представлена цилиндрическими структурами, распространяющимися по всей ширине щели и пронизывающими насквозь оба бислоя. Эти, заполненные водой каналы, служат основой для осуществления физиологических функций щелевого контакта (рис. 7Б ).
Щелевой контакт состоит из субъединиц - коннексонов - с расстоянием между ними 80-100 А, представленных 6 коннексинами полипептидной природы, создающих водный канал между цитоплазмой двух соседних клеток. Одиночные коннексины каждого коннексона могут изгибаться относительно друг друга, открывая или закрывая центральный канал коннексона (рис.7В).
![]() Рис. 7. Межклеточные контакты и структура щелевого контакта.
Рис. 7. Межклеточные контакты и структура щелевого контакта.
А - типы межклеточных контактов. Б - модель структуры щелевого контакта, включающая липидный бислой двух соседних клеток, содержащий коннексоны, каждый из которых построен из шести коннексинов. В - в увеличенном виде представлено открытое и закрытое состояние канала коннексона
Структура коннексонов (рис. 8)
Коннексоны позволяют осуществлять обмен ионами и водорастворимыми молекулами с молекулярной массой до 1200-1500 Да между клетками: возможность метаболической кооперации между клетками, например, АМФ, АДФ или АТФ, цАМФ(вторичный мессенджер) могут проходить через щелевой контакт. Эти каналы служат основой и для электротонического взаимодействия между клетками. Центральный канал коннексона может закрываться в результате увеличения концентрации внутриклеточного Са2+ или Н+ в одной из клеток, а также в ответ на деполяризацию одной или обеих клеток.
В целом, щелевые контакты обладают следующими свойствами и функциями:
Пространственная модель щелевого контакта и молекулярная организация коннексина представлены на рис. 8 А, Б.
![]() Рис. 8. Коннексоны и их молекулярная организация
Рис. 8. Коннексоны и их молекулярная организация
Роль щелевого контакта в проведении возбуждения в ткани
Мембрана электровозбудимых клеток потенциалуправляема, т.е. работа ионных каналов управляется потенциалом, а изменение потенциала зависит от тока, поступающего, например, через щелевой контакт от соседних клеток. При достижении потенциала порога электровозбудимые клетки генерируют потенциалы действия, распространяющиеся, например у нейронов, по отросткам к другим нервным клеткам или эффекторным органам, у мышечных клеток запускают сокращение, а у возбудимых клеток желез управляют секрецией.
Таким образом, функционирование возбудимых тканей связано с распределением потенциала и распространением тока в них.
Транспорт веществ через щелевой контакт
Одно из важнейших свойств щелевого контакта как транспортной системы состоит в том, что при его наличии между соседними клетками может осуществляться интенсивный диффузионный обмен гидрофильными соединениями с молекулярной массой до 1500 Да, минуя наружную среду. Через коннексоны щелевого контакта проходят неорганические ионы (Na+, K+, Cl-, I-, SO42-), сахара (мальтоза, мальтотриоза, мальтотетроза, сахароза), аминокислоты (аспарагиновая, глутаминовая, гексоглицин), нуклеотиды (гипоксантин, аденин, уридин, тиогуанин). Многие из перечисленных веществ не способны диффундировать через наружную мембрану клеток.
Электрический синапс
Механизм передачи сигнала через электрический синапс аналогичен механизму распространения потенциала действия по нервному волокну. В нервном волокне потенциал действия возникает за счет разности потенциалов между возбужденной и невозбужденной областями. Это вызывает открытие Na+-каналов и генерацию импульса заново на каждом последующем участке волокна. В электрическом синапсе в результате разности потенциалов между возбужденным и невозбужденным участками возникают локальные токи, и потенциал из возбужденной терминали аксона распространяется в постсинаптическую клетку, пройдя через ее мембрану и замыкаясь снаружи. Это приводит к открытию Na+-каналов в мембране постсинаптической клетки и возникновению там потенциала действия. Такой механизм работы требует низкого сопротивления пре- и постсинаптической мембран, что обеспечивается наличием коннексонов.
Основные функции электрических синапсов сводятся к следующим.
В сердце электрическая связь показана для клеток всех его отделов, в том числе между клетками с разной дифференцировкой, например между волокнами Пуркинье и клетками рабочего миокарда. Поскольку щелевые контакты между клетками обладают низким сопротивлением относительно мембраны контактирующих клеток, возбужденная клетка может передавать сигнал невозбужденной, в результате чего эта последняя возбуждается.
Нарушение щелевых контактов между клетками ухудшает условия для распространения возбуждения, что может привести к возникновению частичных или полных блоков проведения на отдельных участках ткани и возникновению различных патологических режимов работы сердца. Однако при повреждении или гибели миокардиальных клеток происходит блокирование проводимости коннексонов между нормальной и поврежденной клетками. Это способствует резкой локализации повреждения и увеличивает жизнеспособность сердца.
Лиганд-рецепторное взаимодействие. Лиганд - это молекула, которая может взаимодействовать с участком связывания (местами связывания) молекулярного рецептора - специфическим участком молекулы белковой природы, встроенной, например, в мембрану клетки или находящейся в цитоплазме.
Для реализации механизма связывания необходимо, чтобы лиганд находился практически рядом с участком связывания белка, поскольку в противном случае сила электрического притяжения между противоположно заряженными областями на участке связывания молекулярного рецептора и лиганде будет мала. Лиганд подходит к участку связывания молекулярного рецептора, как ключ к замку.
Взаимодействие лиганда с участком связывания молекулярного рецептора может быть строго специфичным, и, следовательно, участок связывания может связывать только один тип лиганда.
Хотя некоторые участки связывания имеют химическую специфичность, которая позволяет им связывать только один тип лиганда, другие участки менее специфичны и, таким образом, способны связывать ряд родственных лигандов, поскольку фрагмент каждого такого лиганда может быть комплементарен форме участка связывания.
Сила связывания лиганда с участком связывания белка известна как его аффинность или сродство. Мерой сродства участка связывания к лиганду служит концентрация лиганда, необходимая для обеспечения 50% насыщения; чем более низкая концентрация лиганда необходима для связывания его с половиной центров связывания, тем больше афинность участка связывания к лиганду.
Аффинность и химическая специфичность представляют собой два основных свойства участков связывания. Химическая специфичность зависит только от формы участка связывания, а аффинность зависит от силы притяжения между рецептором и лигандом.
Лиганды могут быть агонистами или антагонистами.
Агонист - это лиганд, который взаимодействует с участком связывания и изменяет состояние молекулярного рецептора подобно трансмиттеру.
Наряду с агонистами существуют химические соединения, которые хотя и взаимодействуют с участком связывания на макромолекуле рецептора, но не могут открыть ионный канал. Занимая участок связывания, они препятствуют рецепции естественного трансмиттера и возникновению его эффекта, поскольку действуют на ту же самую макромолекулу рецептора, что и агонист. Такие вещества называют антагонистами.
Химический синапс (рис. 9) выполняет задачу передачи электрического сигнала от пресинаптического нейрона на постсинаптическую клетку. Электрический сигнал пришел на пресинаптическую мембрану и вызвал ее деполяризацию. В результате этого произошло освобождение из везикул пресинаптической зоны специфических переносчиков, называемых нейротрансмиттерами, или медиаторами. Эти химические соединения диффундируют к постсинаптической области и открывают на постсинаптической мембране ионные каналы. Если, например, мембрана клетки станет проницаемой для ионов Na+, то эти ионы по электрохимическому градиенту начнут входить в клетку. В результате происходит деполяризация мембраны клетки. Ионные токи изменяют потенциал мембраны постсинаптической клетки, поэтому такие изменения потенциала называют постсинаптическими потенциалами.
Так как в формировании потенциала покоя мембраны участвует большое число ионов, равновесие может нарушаться посредством изменений проводимости различных ионов. Так, например, при дополнительном выходящем токе ионов K+ или при входящем токе ионов Cl-может происходить гиперполяризация, которая является противоположностью возбуждения, вызывающей торможение нейрона.
Представленные химические процессы могут быть модифицированы посредством других химических веществ. Это происходит при помощи независимых соединений - нейромодуляторов.
Химические процессы в синапсе открывают широкие возможности для фармакологической регуляции и оказываются предметом многочисленных исследований с целью поиска эндогенных соединений, способных модифицировать в заданных направлениях синаптическую передачу.
И действительно, многие медикаменты реализуют свое фармакологическое действие путем влияния на синаптическое проведение. Это относится не только к психотропным и наркотическим веществам. Многие другие, например, (гипотензивные) средства понижающие артериальное давление, также действуют опосредованно через синапсы. Кроме того, многие яды растительного и животного происхождения направленно действуют на химический синапс.
Почти все синаптические терминали освобождают не один только трансмиттер, но одновременно с ним целый ряд биологически активных соединений. Примерами таких сопутствующих соединений служат АТФ, ГТФ, окситоцин, вещество Р, энкефалин и др. Их называют котрансмиттеры.
Многие трансмиттеры, например, глицин и ГАМК или глицин и глутамат могут присутствовать в везикулах одновременно.
Котрансмиттеры могут модулировать синаптические процессы. Накапливаясь в везикулах, они обладают собственной кинетикой освобождения, например освобождение только при значительной величине пресинаптического потенциала.
![]() Рис. 9. Химический синапс и принцип его работы
Рис. 9. Химический синапс и принцип его работы
Механизм высвобождения трансмиттера. Во время деполяризации в нервном окончании возникает не только входящий ток Na+. Мембрана окончания аксона имеет и потенциал-управляемые Ca2+-каналы, через которые во время деполяризации ионы Ca2+ проникают в синаптическое окончание. Одновременно ионы Ca2+ дополнительно выходят из эндоплазматического ретикулума. В синаптическом окончании в зоне пресинаптической мембраны расположено большое число синаптических пузырьков (везикул) с медиатором (трансмиттером).
Преобразование трансмиттера. Трансмиттер может быть либо быстро химически расщеплен на неактивные компоненты, либо удален из синаптической щели путем высокоселективного обратного захвата в пресинаптическое окончание. В центральной нервной системе клетки глии могут также захватывать трансмиттер.
Пресинаптические окончания благодаря описанному механизму обратного захвата (реаптейка) захватывают и возвращают либо фрагменты трансмиттера (например, холин), либо всю молекулу трансмиттера (например, серотонин). Для этого в пресинаптической мембране находятся специфические протеины - транспортеры.
Во многих синапсах рецепторы для трансмиттера есть не только в постсинаптической мембране, но также и в пресинаптической. Это так называемые ауторецепторы.
![]() Рис. 10. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Рис. 10. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Типы химических синапсов.
У части синапсов ионный канал, который при действии лиганда, т.е. благодаря связыванию с молекулой трансмиттера, открывается (каналы, управляемые лигандами), называется ионотропным рецептором (рис. 11 А).
Рецепторы, в которых открытие ионного канала связано с подключением других химических процессов, называют метаботропными рецепторами. На метаботропных рецепторах лиганд открывает ионный канал не прямо, а опосредованно, через вторичные мессенджеры (рис.11Б).
Примером и ионотропного, и метаботропного синапса служит холинергический синапс, у которого трансмиттером служит АЦХ. У холинергических синапсов два типа рецепторов: АЦХ-рецепторы, чувствительные дополнительно к никотину (никотиновый АЦХ-рецептор), и АЦХ-рецепторы, чувствительные дополнительно к мускарину (мускариновый АЦХ-рецептор). Первый является ионотропным, второй - метаботропным.
![]() Рис. 11. Ионотропные и метаботропные синапсы
Рис. 11. Ионотропные и метаботропные синапсы
![]() Рис. 12. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б)
Рис. 12. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б)
Тест
1. В ЦНС человека ключевую роль в передаче информации между клектами играют
a) электрические и химические синапсы
b) химические синапсы
2. Основу электрического синапса у человека играют
a) щелевые контакты
b) десмосомы и щелевые контакты
3. В структуре химического синапса переача сигнала осуществляется посредством
a) нейромедиатора
b) коннексинов
4. Химические синапсы ЦНС представлены следующими типами
a) метаботропными и ионотропными
b) только ионотропными
5. В основе работы метаботропного синапса лежит
a) механизм вторичных мессенджеров
b) изменение проницаемости ионных каналов
Ацетилхолин. Является медиатором для всех двигательных нейронов, многих интернейронов ЦНС, а также нейронов парасимпатической нервной системы. Существует 2 типа рецепторов ацетилхолина: АЦХ-рецепторы, чувствительные к никотину: никотиновый АЦХ-рецептор и, соответственно, никотиновый АЦХ (нАЦХ) синапс, и АЦХ-рецепторы, чувствительные к мускарину - мускариновый АЦХ-рецептор и, соответственно, мускариновый (мАЦХ) АЦХ синапс. АЦХ синапс с никотиновым рецептором являются ионотропными, а АЦХ синапсы с мускариновым рецептором являются метаботропными.
Глутамат в синапсах выполняет функцию трансмиттера и ко-трансмиттера (может присутствовать одновременно с другими трансмиттерами, например с глицином). Глутамат в мозге является важнейшим трансмиттером для возбуждающих синапсов. Синапсы, использующие в качестве трансмиттера глутамат, находятся приблизительно на 50% нейронов центральной нервной системы. Такие синапсы образуют важнейшие возбуждающие входы систем мозга к коре большого мозга. Они принимают участие в процессах обучения. Глутамат прямо открывает неспецифический ионный канал для катионов.
Глицин служит трансмиттером тормозных синапсов и действует как нейромодулятор. Глицин предназначен для выполнения специфических ингибиторных задач. Большинство клеток Реншоу, через которые тормозятся α-мотонейроны спинного мозга, выбрасывают глицин. Глициновые рецепторы открывают каналы для ионов Cl-, что способствует возникновению так называемых тормозных постсинаптических потенциалов (тормозных потенциалов синаптической пластинки - ТПСП). Синаптическое действие заканчивается с помощью механизма обратного захвата глицина.
В центральной нервной системе глицин играет также и роль нейромодулятора, например, в гиппокампе NMDA-рецепторы, описанные в предыдущем разделе, испытывают модулирующее влияние со стороны глицина.
γ-Аминомасляную кислоту (ГАМК) выбрасывают в качестве трансмиттера многие тормозные интернейроны, имеющиеся практически во всех отделах центральной нервной системы, и аксоны клеток Пуркинье мозжечка. ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей мере два различных постсинаптических механизма, которые запускаются благодаря различным рецепторам (ГАМКА и ГАМКС с одной стороны; ГАМКВ с другой стороны).
Серотонин (5-hydroxytryptamin) - биогенный амин, широко распространенный в ЦНС. В мозге его особенно много в области тектума. Оттуда наблюдается много проекций в лимбическую систему, к таламусу и гипоталамусу, в передний мозг, в мозжечок и в спинной мозг. По этим путям, очевидно, осуществляется контроль многих нейрональных функций. На постсинаптической мембране были обнаружены различные серотониновые рецепторы, которые большей частью, благодаря вторичным мессенджерам, открывают каналы для ионов К+ и Са2+. Действие трансмиттера заканчивается включением механизма обратного захвата пресинаптической областью.
Гистамин - моноамин, выступающий в качестве трансмиттера. Особенно важную роль он играет как модулятор в мозге грудных детей. Гистаминергические нейроны у взрослых находятся в заднем гипоталамусе и связаны со многими участками мозга, где они оказывают влияние на состояние бодрствования, мышечную активность, прием пищи, сексуальные отношения и обменные процессы в мозге. Из-за участия этих нейронов в регуляции процессов сна и бодрствования многие антигистаминные препараты вызывают состояние сонливости.
Дофамин - биогенный амин, наиболее представленный в базальных ганглиях, где нейроны черной субстанции образуют дофаминергический путь к стриатуму, выполняя функцию контроля моторики.
Дофаминергические пути, которые исходят от area ventalis tegmentalis и проецируются к nucleus accumbeus,играют большую роль в патогенезе эпилепсии.
Существуют различные дофаминовые рецепторы, действующие через вторичные мессенджеры. Их постсинаптическое действие может быть тормозным или возбуждающим. Дофамин очень быстро захватывается обратно из синаптической щели в пресинаптическое окончание. Там он претерпевает превращения благодаря моноаминоксидазе. Вне нейрона, он подвергается воздействию катехол-О-метилтрансферазы.
Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС и синтезирующийся прежде всего нейронами locus coeruleus. Это ядро состоит не более чем из 1000 клеток, аксоны которых так многократно разветвляются, что можно найти адренергические окончания во многих областях ЦНС. Они оказывают модулирующее влияние, например, на процессы созревания, обучения, переработку информации в мозгу, регуляцию сна и на эндогенное торможение боли. В периферической нервной системе норадреналин служит важным трансмиттером симпатических постганглионарных окончаний, например, на сердце и на гладких мышцах сосудов. В некоторых центральных синапсах как трансмиттер рассматривают адреналин.
Для катехоламинов существует четыре главных типа рецепторов: α1, α2, β1 и β2. Они отличаются по реакции на различные агонисты или антагонисты, а также по постсинаптическим эффектам. Рецепторы α1 управляют Са2+-каналами при помощи вторичного мессенджера IP3 и при активации повышают внутриклеточную концентрацию ионов Са2+. Активация рецепторов ведет к уменьшению концентрации вторичного мессенджера цАМФ, вызывая различные эффекты. Активация рецепторов β, например, на клетках Пуркинье мозжечка, повышает при помощи вторичного мессенджера цАМФ проводимость мембраны для ионов К+ и, таким образом, образует ТПСП. Обратный захват и разрушение похожи на таковые для дофамина.
Олигопептиды, которые действуют как трансмиттеры или нейромодуляторы - это приемущественно короткие полипептиды, состоящие из 2-30 аминокислотных остатков. Они функционируют как трансмиттеры или ко-трансмиттеры либо модулируют синаптические процессы. Так энкефалин, эндорфин и динорфин служат трансмиттерами в тех синапсах, в которых опиоидные вещества также оказывают действие. Опиоиды (например, морфин) являются сильными анальгетиками. Вышеназванные опиоидные пептиды тормозят распространение боли в спинном мозге. Кроме того, они играют большую роль в лимбической, автономной и моторной системах.
Другие нейропептиды: вещество Р, ангиотензин II, соматостатин, вазоактивный интестинальный полипептид (VIP), нейропептид Y и многие другие. Большинство названных веществ были открыты в качестве гормонов прежде, чем была определена их роль в синаптическом переносе.
ГАМК как трансмиттер
Многие тормозные интернейроны, присутствующие практически во всех отделах ЦНС, и аксоны клеток Пуркинье мозжечка выбрасывают в качестве трансмиттера γ-аминомасляную кислоту (ГАМК, GABA: γ-Aminobutyric acid). ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей мере два различных постсинаптических механизма, запускающихся благодаря различным рецепторам (ГАМКА, ГАМКС с одной стороны; ГАМКВ с другой стороны).
ГАМКА-и ГАМКС-рецепторы непосредственно открывают каналы для ионов Cl-, что приводит к появлению входящего Cl-тока и тем самым к гиперполяризации в виде ТПСП. Барбитураты (препараты, применяемые для наркоза), стероидные анестетики и транквилизаторы (успокаивающие средства) из класса бензодиазепинов (например, диазепам) усиливают ингибиторное действие ГАМКА-рецепторов. Механизм их действия заключается в связывании с белком ионного канала, который уже взаимодействует с ГАМК, и таким образом в усилении действия ГАМК.
Канал построен из нескольких субъединиц. Одна из этих субъединиц связывает ГАМК, другая может дополнительно связать бензодиазепин, третья - связать барбитурат и еще одна - связать стероид. В настоящее время неясно, какие природные вещества (например, нейромодуляторы) оказывают действие на так называемые бензодиазепиновые или барбитуратные рецепторы. Названные субъединицы могут обладать различными химическими свойствами, определяемыми их химической структурой, что обусловливает различные фармакологические эффекты. БикукуллинÄ представляет собой конкурентный блокатор для ГАМКА-, но не для ГАМКС-рецептора. Яд пикротоксин, вызывающий судороги, является неконкурентным блокатором, закрывающим канал для ионов Cl-.
ГАМКВ открывают каналы для ионов К+ посредством G-белка. Агонистом служит баклофен. Пресинаптические ГАМКВ-рецепторы закрывают каналы Са2+и тормозят освобождение трансмиттера. Действие ГАМК прекращается путем обратного захвата ГАМК пресинаптическим окончанием и клетками глии.
![]() Рис. 13. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный захват
Рис. 13. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный захват
Глицин как трансмиттер
Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует как нейромодулятор. Глицин выполняет специфические ингибиторные задачи. Большинство клеток Реншоу, через которые осуществляется торможение α-мотонейронов спинного мозга, высвобождают глицин. Стрихнин является конкурентным антагонистом, и его применение приводит к судорогам из-за отсутствия торможения α-мотонейронов. Рецепторы глицина открывают С1--каналы, что способствует возникновению ТПСП. Синаптическое действие заканчивается с помощью механизма обратного захвата глицина.
В ЦНС глицин играет также и роль нейромодулятора, например в гиппокампе NMDA-рецепторы, описанные ранее, испытывают модулирующее влияние со стороны глицина.
Важно заметить, что хотя синапсы называются по наименованию их главного трансмиттера (например, холинергический).
NO как трансмиттер
Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками и некоторыми нейронами. Вследствие того, что NO быстро окисляется, его биологическая жизнь длится всего несколько секунд. По этой причине NO влияет только на клетки, находящиеся вблизи от места своего выделения. NO стимулирует растворимую гуанилатциклазу в клетках-мишенях и тем самым повышает в клетке мишени внутриклеточную концентрацию цГМФ. Это, в свою очередь, стимулирует цГМФ-зависимую протеинкиназу.
Оксид азота легко распространяется путем диффузии и может принимать на себя функции трансмиттера. При этом, например, пресинаптические процессы могут регулироваться с помощью обратной связи. Возможно, этот вид переноса информации играет большую роль в ЦНС. Механизмы прекращения действия подобных трансмиттеров не позволяют включить их в классическую синаптическую схему.
Тест
1. Что из перечисленного не является нейротрансмиттером?
a) ацетилхолин, норадреналин, ГАМК, серотонин
b) холинэстераза, катехол-О-метилтрасфераза
2. Верно ли следующее утверждение. Оксид азота не является нейротрансмиттером т.к. это вещество очень быстро окисляется
a) да, верно
b) нет, не верно
3. Верно ли утверждение, что каждый нейротрансмиттер может связываться только с ионотропным или только с метаботропным рецептором?
a) да, верно
b) нет, не верно
5. Постсинаптические потенциалы
Схема распространения информации по нейрону (рис.14)
В зоне синапса локальный возбуждающий постсинаптический потенциал (ВПСП) распространяется пассивно электротонически по всей постсинаптической мембране клетки. Это распространение не подчиняется закону «все или ничего». Если большое число возбуждающих синапсов возбуждаются одновременно или почти одновременно, то возникает явление суммации, проявляющееся в виде возникновения ВПСП существенно большей амплитуды, что может деполяризовать мембрану всей постсинаптической клетки. Если величина этой деполяризации достигает в области постсинаптической мембраны порогового значения (10 мВ или больше), то на аксонном холмике нервной клетки молниеносно открываются потенциалуправляемые Na+-каналы, и клетка генерирует потенциал действия, проводящийся вдоль ее аксона. От начала ВПСП до образования потенциала действия проходит еще около 0,3 мс.
Пороговый стимул - самый слабый стимул, надежно различаемый сенсорным рецептором. Для этого стимул должен вызывать рецепторный потенциал такой амплитуды, которая достаточна для активации хотя бы одного первичного афферентного волокна. Более слабые стимулы могут вызвать подпороговый рецепторный потенциал, однако они не приведут к возбуждению центральных сенсорных нейронов и, следовательно, не будут восприняты. Кроме того, количество возбужденных первичных афферентных нейронов, необходимое для сенсорного восприятия, зависит от пространственной и временной суммации в сенсорных путях (рис. 14, Г).
Взаимодействуя с рецептором, молекулы АЦХ открывают неспецифические ионные каналы в постсинаптической мембране клетки так, что повышается их способность к проводимости одновалентных катионов. Работа каналов ведет к базовому входящему току положительных ионов, и, следовательно, к деполяризации постсинаптической мембраны, которая применительно к синапсам называется возбуждающим постсинаптическим потенциалом.
Ионные токи, участвующие в возникновении ВПСП, ведут себя иначе, нежели токи натрия и калия во время генерации потенциала действия. Причина заключается в том, что в механизме генерации ВПСП участвуют другие ионные каналы с другими свойствами (лигандуправляемые, а не потенциалуправляемые). При потенциале действия активируются потенциалуправляемые ионные каналы, и с увеличивающейся деполяризацией открываются следующие каналы, так что процесс деполяризации усиливает сам себя. В то же время проводимость трансмиттеруправляемых каналов (лигандуправляемых) зависит только от количества молекул трансмиттера, связавшихся с молекулами рецептора (в результате чего открываются трансмиттеруправляемые ионные каналы) и, следовательно, от числа открытых ионных каналов. Амплитуда ВПСП лежит в диапазоне от 100 μВ до в некоторых случаях 10 мВ. В зависимости от вида синапса, общая продолжительность ВПСП у некоторых синапсов находится в диапазоне от 5 до 100 мс.
![]() Рис. 14. Информация течет от дендритов к соме, к аксону, к синапсу. На рисунке представлены типы потенциалов в разных местах нейрона в зависимости от пространственной и временной суммации
Рис. 14. Информация течет от дендритов к соме, к аксону, к синапсу. На рисунке представлены типы потенциалов в разных местах нейрона в зависимости от пространственной и временной суммации
Возбуждающий постсинаптический потенциал (ВПСП)
Рассмотрим этот вопрос на примере АЦХ. Взаимодействуя с рецептором, молекулы АЦХ открывают неспецифические ионные каналы в постсинаптической мембране клетки так, что повышается их способность к проводимости для одновалентных катионов. Существует сильная движущая сила для натрия, и ионы Na+устремляются внутрь клетки и деполяризуют ее мембрану. С другой стороны, канал проходим и для ионов K+, для которых сохраняется незначительный электрохимический градиент, направленный из внутриклеточной области к внеклеточной среде. Через постсинаптическую мембрану одновременно проходят и ионы K+, тем самым слегка противодействуя деполяризации, обусловленной входящим током ионов Na+. Работа описанных каналов ведет к базовому входящему току положительных ионов и, следовательно, к деполяризации постсинаптической мембраны, которая называется применительно к синапсам ВПСП. На концевой пластинке нервно-мышечного синапса ВПСП называют также потенциалом концевой пластинки. Так как участвующие ионные токи зависят от разности их потенциала равновесия и потенциала мембраны, то при уменьшенном потенциале покоя мембраны ток ионов Na+ становится всегда меньше, а ток ионов К+- больше. Именно поэтому амплитуда ВПСП уменьшается.
В механизме генерации ВПСП участвуют и другие ионные каналы с другими свойствами (лигандуправляемые, а не потенциал-управляемые), и проводимость трансмиттер-управляемых каналов (лиганд-управляемых) зависит только от количества молекул трансмиттера, связавшихся с молекулами рецептора. Амплитуда ВПСП лежит в диапазоне от 100 μВ до (в некоторых случаях) 10 мВ. В зависимости от вида синапса, общая продолжительность ВПСП лежит в диапазоне от 5 до 100 мс.
Прежде всего, в зоне синапса образовавшийся локально ВПСП распространяется пассивно электротонически по всей постсинаптической мембране клетки. Этот процесс не подчиняется закону «все или ничего». Если большое число возбуждающих синапсов активируются одновременно или почти одновременно, то возникает явление так называемой суммации, которое реализуется в виде возникновения ВПСП существенно большей амплитуды, что может деполяризовать мембрану всей постсинаптической клетки. Если величина этой деполяризации достигает в области постсинаптической мембраны порогового значения (10 мВ или больше), то на аксонном холмике нервной клетки молниеносно открываются потенциал-управляемые Na+-каналы, и клетка генерирует потенциал действия, который проводится вдоль аксона. В случае моторной концевой пластинки это приводит к мышечному сокращению. От начала ВПСП до образования потенциала действия проходит еще около 0,3 мс, так что при обильном освобождении трансмиттера постсинаптический потенциал может появиться уже через 0,5-0,6 мс после пришедшего в пресинаптическую область потенциала действия. В общих чертах, время «синаптической задержки», подразумевающее необходимое время между возникновением пре- и постсинаптического потенциала действия, всегда зависит от типа синапса.
Тормозной постсинаптический потенциал (ТПСП)
Решающими для ионного тока в мембране оказываются величина ее потенциала и количество открытых ионных каналов. Определяющим оказывается тип белка канала, на который действует трансмиттер. Так на одних синапсах находятся каналы для K+, тогда как на других есть каналы для Cl-. Последние распространены много чаще. Рассмотрим в виде примера рецептор метаботропного синапса, повышающий при связывании с трансмиттером проводимость для ионов К+. При нормальной величине потенциала мембраны это приводит к дальнейшему выходящему току ионов К+ и гиперполяризации потенциала мембраны. Возникает так называемый ТПСП. Принципиально похожая ситуация складывается, если гиперполяризующий мембрану ток связан с ионами Cl-. Cl- течет в клетку и гиперполяризует ее. Подобная картина характерна для очень многих клеток.
![]() Рис. 15. Потенциалы и токи, возникающие на постсинаптической мембране.
Рис. 15. Потенциалы и токи, возникающие на постсинаптической мембране.
А - процессы освобождения трансмиттера и формирование возбуждающего постсинаптического потенциала (ВПСП) после электрического возбуждения (стрелка), поступившего на пресинаптическое окончание, связаны с возникновением возбуждающего постсинаптического тока (ВПСТ) и генерацией вследствие этого ВПСП. Если ВПСП переходит порог, то возникает потенциал действия (ПД), который во время фазы нарастания благодаря потенциалуправляемым Na+каналам деполяризует мембрану в направлении ENa, т.е. к потенциалу равновесия для Na+. Б - процессы освобождения трансмиттера и формирование ТПСП связаны с ингибиторным постсинаптическим током (ТПСТ) и соответствующим ему ТПСП. При этом ток вызывает движение через мембрану ионов К+. Ионы Cl- могут играть роль, если потенциал мембраны позитивнее, чем потенциал равновесия для Cl-, который лежит от -75 до -70 мВ. В - токи мембраны при активации возбуждающих (ВПСТ) и ингибиторных синапсов (ТПСТ) и возникающие в этом случае результирующие постсинаптические потенциалы (ВПСП, ТПСП). При одновременной активации возбуждающих и тормозных синапсов возникающие мембранные токи суммируются, поэтому результирующий постсинаптический потенциал (красный) становится очень маленьким
Взаимное влияние ВПСП и ТПСП
Если на мембране клетки активируются одновременно возбуждающие и тормозные синапсы, то ионный ток уменьшается.
Если соседствующие возбуждающие и тормозные синапсы (одновременно) активируются, то возникающие токи накладываются друг на друга. Результирующий постсинаптический потенциал (по абсолютной величине) меньше, чем только один ВПСП или только один ТПСП.
ВПСП возникает благодаря повышению проводимости для натрия и тем самым благодаря входящему току натрия, а ТПСП - благодаря выходящему току калия или входящему току хлора.
Тест
1. На постсинаптической мембране могут возникать как возбуждающие, так и тормозные потенциалы, что зависит от
a) типа медиатора и рецепторов, открывающих разные ионные каналы
b) только от типа медиатора
2. Возбуждающий постсинаптический потенциал связан с открытием
a) натриевых и кальциевых ионных каналов
b) хлорных и калиевых ионных каналов
3. Тормозный постсинаптический потенциал связан с открытием
a) натриевых и кальциевых ионных каналов
b) хлорных и калиевых ионных каналов
4. ВПСП – это всегда
a) деполяризация мембраны
b) гиперполяризация мембраны
5. ТПСП – это всегда
a) гиперполяризация мембраны
b) деполяризация мембраны
Существуют несколько типов аксонального транспорта. Окруженные мембранами органоиды и митохондрии транспортируются с относительно большой скоростью посредством быстрого аксонального транспорта. Вещества, растворенные в цитоплазме (например, белки), перемещаются с помощью медленного аксонального транспорта. У млекопитающих быстрый аксональный транспорт имеет скорость 400 мм/сутки, а медленный аксональный транспорт - около 1 мм/сутки.
Для аксонального транспорта требуется затрата метаболической энергии и присутствие внутриклеточного Са2+. Элементы цитоскелета (точнее, микротрубочки) создают систему направляющих тяжей, вдоль которых передвигаются окруженные мембранами органоиды.
Аксональный транспорт осуществляется в двух направлениях. Транспорт от сомы к аксональным терминалям, называемый антероградным аксональным транспортом, восполняет в пресинаптических окончаниях запас синаптических пузырьков и ферментов, ответственных за синтез нейромедиатора. Транспорт в противоположном направлении - ретроградный аксональный транспорт, возвращает опустошенные синаптические пузырьки в сому, где эти мембранные структуры деградируются лизосомами.
Высокая, в ряде случаев, потребность в молекулах трансмиттера вызывает, как правило, синтез трансмиттера на месте, то есть непосредственно в пресинаптическом окончании.
Теперь необходимо отметить, что хотя синапсы называются на основании названия их главного трансмиттера (например, холинэргический), почти все синаптические терминали освобождают не только один единственный трансмиттер, но одновременно с ним целый ряд биологически активных соединений - ко-трансмиттеров. Примерами таких сопутствующих соединений являются АТФ, ГТФ, окситоцин, вещество Р, энкефалин, и другие. Ко-трансмиттеры могут модулировать синаптический процесс.
![]() Рис. 16. Синтез и рецикл синаптических везикул и их содержимого.
Рис. 16. Синтез и рецикл синаптических везикул и их содержимого.
А - Биосинтез нейротрансмиттеров.
I. Везикулы, предшественники пептидных трансмиттеров и ферменты синтезируются в теле нейрона и высвобождаются из аппарата Гольджи.
II. Везикулы перемещаются по аксону с помощью быстрого аксонального транспорта. Пептидные трансмиттеры уже заключены в некоторые везикулы.
III. Непептидные нейротрансмиттеры синтезируются и транспортируются в везикулы непосредственно в нервных окончаниях
Б - Экзоцитоз (высвобождение визикул с медиатором)
Тест
1. Органоиды перемещаются вдоль аксона путем
а) быстрого аксонального транспорта
б) медленного аксонального транспорта
2. Молекулы растворенных в цитоплазме нейрона веществ перемещаются вдоль аксона путем
а) медленного аксонального транспорта
б) быстрого аксонального транспорта
3. Антероградный аксональный транспорта имеет направление
а) от сомы к аксональным терминалям
б) от нервного окончания к соме
4. Ретроградный аксональный транспорт имеет направление
а) от сомы к аксональным терминалям
б) от нервного окончания к соме
Общий объем ЦСЖ в желудочках мозга - примерно 35 мл, тогда как подпаутинное пространство содержит около 100 мл. Каждую минуту образуется примерно 0,35 мл ЦСЖ. При такой скорости обновление ЦСЖ происходит приблизительно четыре раза в сутки.
У человека в положении лежа давление ЦСЖ в спинно-мозговом субарахноидальном пространстве достигает 120-180 мм вод.ст. Скорость образования ЦСЖ относительно независима от давления в желудочках и в субарахноидальном пространстве, а также от системного кровяного давления. В то же время скорость обратного всасывания ЦСЖ прямо связана с давлением ЦСЖ.
Внеклеточная жидкость в ЦНС непосредственно сообщается с ЦСЖ. Следовательно, состав ЦСЖ влияет на состав внеклеточной среды вокруг нейронов головного и спинного мозга. Основные компоненты ЦСЖ в поясничной цистерне перечислены в табл. 5-1. Для сравнения приведены концентрации соответствующих веществ в крови. Как показано в данной таблице, содержание К+, глюкозы и белков в ЦСЖ ниже, чем в крови, а содержание Na+ и Cl- - выше. Кроме того, в ЦСЖ практически нет эритроцитов. Благодаря повышенному содержанию Na+ и Cl- обеспечивается изотоничность ЦСЖ и крови, несмотря на то, что в ЦСЖ относительно мало белков.
Таблица. Состав цереброспинальной жидкости и крови
![]()
План лекции
- Общая характеристика функций нервной системы.
- Нейрон как структурно-функциональная единица НС. Нейрон - возбудимая клетка. Глиальные клетки.
- Физиология синапсов. Типы. Строение. Механизмы передачи возбуждения.
- Нейромедиаторы. Нейромодуляторы.
- Постсинаптические потенциалы.
- Аксональный транспорт.
- Гематоэнцефалический барьер и цереброспинальная жидкость.
- 1. Общая характеристика функций нервной системы
Центральная нервная система (ЦНС)
- собирает и перерабатывает поступающую от периферической нервной системы (ПНС) информацию об окружающей среде, формирует рефлексы и другие поведенческие реакции, планирует (подготавливает) и осуществляет произвольные движения,
- обеспечивает высшие познавательные (когнитивные) функции,
- обеспечивает процессы, связанные с памятью, обучаемостью и мышлением,
- осуществляет координацию деятельности и управляет функциями всех вну ренних органов и оменных процессов.
- спинной мозг подразделяется на последовательные отделы (шейный, грудной, поясничный, крестцовый и копчиковый), каждый из которых состоит из сегментов,
- головной мозг подразделяют на пять отделов: myelencephalon (продолговатый мозг), metencephalon (задний мозг), mesencephalon (средний мозг), diencephalon (промежуточный мозг) и telencephalon (конечный мозг),
- кора каждого полушария головного мозга состоит из долей, которые названы так же, как соответствующие кости черепа: лобная (lobus frontalis), теменная (l. parietalis), височная (l. temporalis) и затылочная (l. occipitalis) доли; полушария соединены мозолистым телом (corpus callosum) - массивным пучком аксонов, пересекающих среднюю линию между полушариями,
- на поверхности ЦНС лежат несколько слоев соединительной ткани - мозговые оболочки: мягкая (pia mater), паутинная (arachnoidea mater) и твердая (dura mater). Они защищают ЦНС. Подпаутинное (субарахноидальное) пространство между мягкой и паутинной оболочками заполнено цереброспинальной (спинно-мозговой) жидкостью (ЦСЖ).
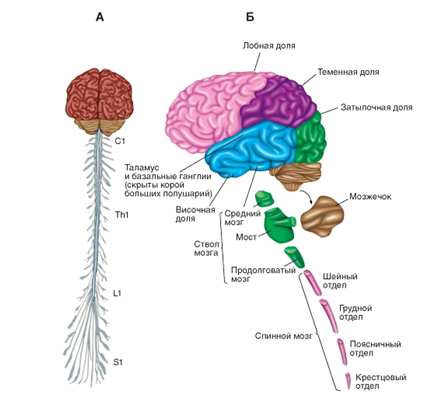 Рис.1. Строение центральной нервной системы.
Рис.1. Строение центральной нервной системы. А -головной и спинной мозг со спинальными нервами. Б - основные компоненты центральной нервной системы. Показаны также четыре главные доли коры больших полушарий: затылочная, теменная, лобная и височная
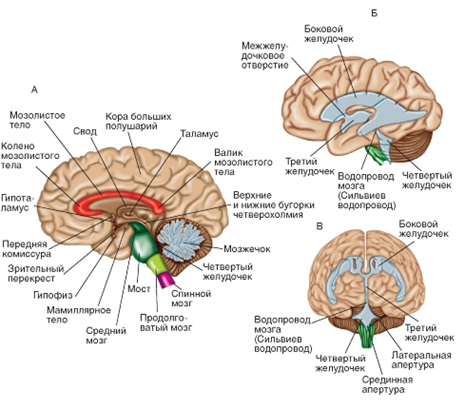 Рис. 2. Головной мозг. А - среднесагиттальный срез головного мозга. Обратите внимание на относительное расположение коры больших полушарий, мозжечка, таламуса и ствола мозга, а также различных комиссур. Б и В - система желудочков мозга in situ - вид сбоку (Б) и спереди (В)
Рис. 2. Головной мозг. А - среднесагиттальный срез головного мозга. Обратите внимание на относительное расположение коры больших полушарий, мозжечка, таламуса и ствола мозга, а также различных комиссур. Б и В - система желудочков мозга in situ - вид сбоку (Б) и спереди (В) Тест
1. ЦНС представлена
a) спинным и головным мозгом
b) головным мозгом
2. Выберите наиболее правильный ответ. Основная роль ЦНМ заключается в
a) переработке информации от ПНС и выработке программы действия
b) регуляции движением
- Нейрон как структурно-функциональная единица НС. Нейрон - возбудимая клетка. Глиальные клетки.
Функциональная единица нервной системы – нейрон (рис 3).
Типичный нейрон обладает воспринимающей поверхностью в виде тела (сомы) и нескольких отростков – дендритов, на которых находятся синапсы, т.е. межнейронные контакты. Аксон нервной клетки образует синаптические связи с другими нейронами или с эффекторными клетками. Коммуникативные сети нервной системы складываются из нейронных цепей, образованных синаптически взаимосвязанными нейронами.
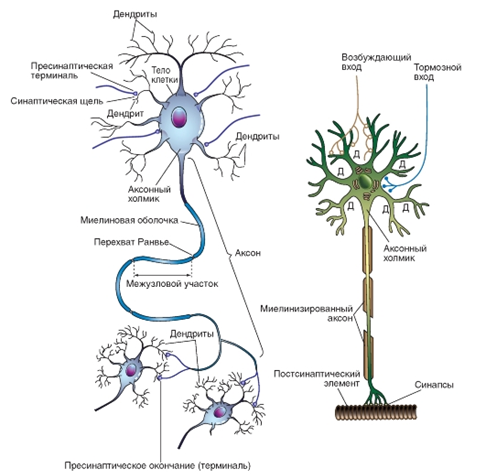 Рис. 3. Схема «идеального» нейрона и его основных компонентов.
Рис. 3. Схема «идеального» нейрона и его основных компонентов. Большинство афферентных входов, поступающих по аксонам других клеток, оканчиваются синапсами на дендритах (Д), но некоторые - синапсами на соме. Возбуждающие нервные окончания чаще располагаются дистально на дендритах, а тормозные нервные окончания чаще находятся на соме.
Сома. В соме нейронов находятся ядро и ядрышко (рис. 3), а также хорошо развитый аппарат биосинтеза, который производит компоненты мембран, синтезирует ферменты и другие химические соединения, необходимые для специализированных функций нервных клеток. К аппарату биосинтеза в нейронах относятся тельца Ниссля - плотно прилегающие друг к другу сплюснутые цистерны гранулярного эндоплазматического ретикулума, а также хорошо выраженный аппарат Гольджи. Кроме того, сома содержит многочисленные митохондрии и элементы цитоскелета, в том числе нейрофиламенты и микротрубочки. В результате неполной деградации мембранных компонентов образуется пигмент липофусцин, накапливающийся с возрастом в ряде нейронов. В некоторых группах нейронов ствола мозга (например, в нейронах черной субстанции и голубого пятна) заметен пигмент мелатонин.
| |
Аксон. Специализированный участок тела клетки (чаще сомы, но иногда - дендрита), от которого отходит аксон, называется аксонным холмиком. Аксон и аксонный холмик отличаются от сомы и проксимальных участков дендритов тем, что в них нет гранулярного эндоплазматического ретикулума, свободных рибосом и аппарата Гольджи. В аксоне присутствуют гладкий эндоплазматический ретикулум и выраженный цитоскелет.
Нейроны сообщаются друг с другом с помощью потенциалов действия, распространяющихся в нейронных цепях по аксонам. Потенциалы действия поступают от одного нейрона к следующему в результате синаптической передачи. В процессе передачи достигший пресинаптического окончания потенциал действия обычно запускает высвобождение нейромедиатора, который либо возбуждает постсинаптическую клетку, так что в ней возникает потенциал действия, либо тормозит ее активность. Аксоны не только передают информацию в нейронных цепях, но и доставляют путем аксонального транспорта химические вещества к синаптическим окончаниям.
Органеллы нейрона. На рисунке 4 представлена сома нейронов. В соме нейронов показаны ядро и ядрышко, аппарат биосинтеза, который производит компоненты мембран, синтезирует ферменты и другие химические соединения, необходимые для специализированных функций нервных клеток. В него входят тельца Ниссля - цистерны гранулярного эндоплазматического ретикулума, а также хорошо выраженный аппарат Гольджи. Сома содержит митохондрии и элементы цитоскелета, в том числе нейрофиламенты и микротрубочки. В результате неполной деградации мембранных компонентов образуется пигмент липофусцин, накапливающийся с возрастом в ряде нейронов. В некоторых группах нейронов ствола мозга (например, в нейронах черной субстанции и голубого пятна) заметен пигмент мелатонин.
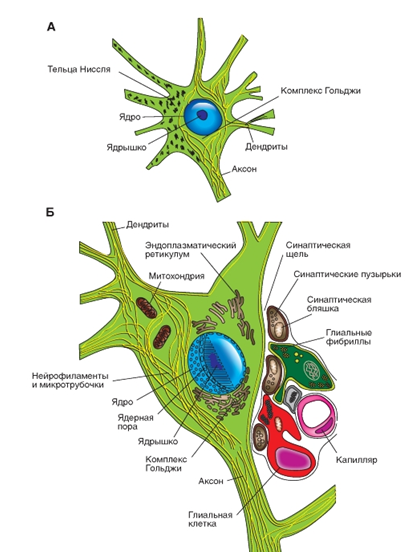 Рис. 4. Нейрон. А - органеллы нейрона. На поверхности нейрона - синаптические окончания. Б - Схема соответствует электронно-микроскопической картине. Видны ядро, ядрышко, хроматин, ядерные поры. В цитоплазме - митохондрии, шероховатый эндоплазматический ретикулум, аппарат Гольджи, нейрофиламенты и микротрубочки. На наружной стороне плазматической мембраны - синаптические окончания и отростки астроцитов.
Рис. 4. Нейрон. А - органеллы нейрона. На поверхности нейрона - синаптические окончания. Б - Схема соответствует электронно-микроскопической картине. Видны ядро, ядрышко, хроматин, ядерные поры. В цитоплазме - митохондрии, шероховатый эндоплазматический ретикулум, аппарат Гольджи, нейрофиламенты и микротрубочки. На наружной стороне плазматической мембраны - синаптические окончания и отростки астроцитов. Типы нейронов
Нейроны разного типа выполняют специфичные коммуникативные функции (рис. 5).
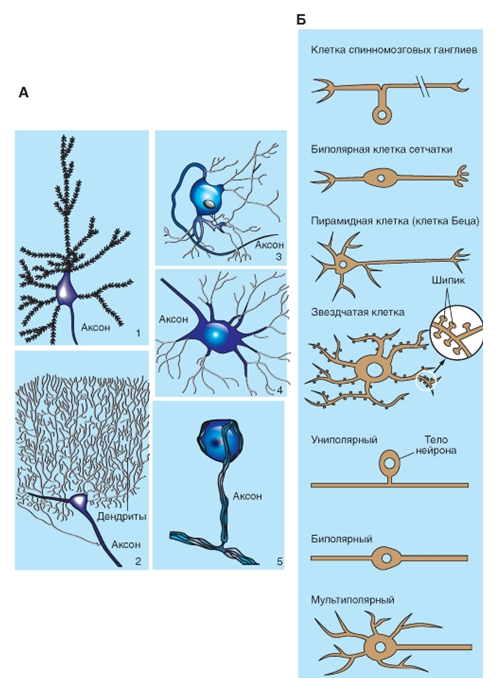 Рис. 5. Типы нейронов
Рис. 5. Типы нейронов А - нейроны разнообразной формы: 1 - нейрон, напоминающий пирамиду (пирамидные клетки коры больших полушарий); 2 - клетки Пуркинье в коре мозжечка; 3 - постганглионарный симпатический мотонейрон; 4 - альфа-мотонейрон спинного мозга, мультиполярный, с радиальными дендритами; 5 - сенсорная клетка спинального ганглия; не имеет дендритов, ее отросток разделяется на две ветви: центральную и периферическую, псевдоуниполярные нейроны. Б - типы нейронов
Так, нейроны ганглиев задних корешков (спинальных ганглиев) получают информацию от сенсорных нервных окончаний в органах. Их клеточные тела лишены дендритов (рис. 5 ) и не получают синаптических окончаний. Выйдя из клеточного тела, аксон такого нейрона разделяется на две ветви, одна из которых (периферический отросток) направляется в составе периферического нерва к сенсорному рецептору, а другая ветвь (центральный отросток) входит в спинной мозг (в составе заднего корешка) либо в ствол мозга (в составе черепного нерва).
Нейроны другого типа, такие, как пирамидные клетки коры больших полушарий и клетки Пуркинье коры мозжечка, заняты переработкой информации (рис. 5). Их дендриты покрыты дендритными шипиками и характеризуются обширной поверхностью. Они имеют огромное количество синаптических входов.
Виды ненейронных клеток (нейроглия)
Нейроглии (рис. 6) в ЦНС человека на порядок больше, чем число нейронов: 1013 и 1012 соответственно. Нейроглия способствует осуществлению коммуникативных процессов нейронами, образует миелиновую оболочку, увеличивающую скорость проведения потенциалов действия.
Типы нейроглии
В ЦНС к нейроглии относят астроциты и олигодендроциты, клетки микроглии и эпендимы, а в ПНС - шванновские клетки и клетки-сателлиты.
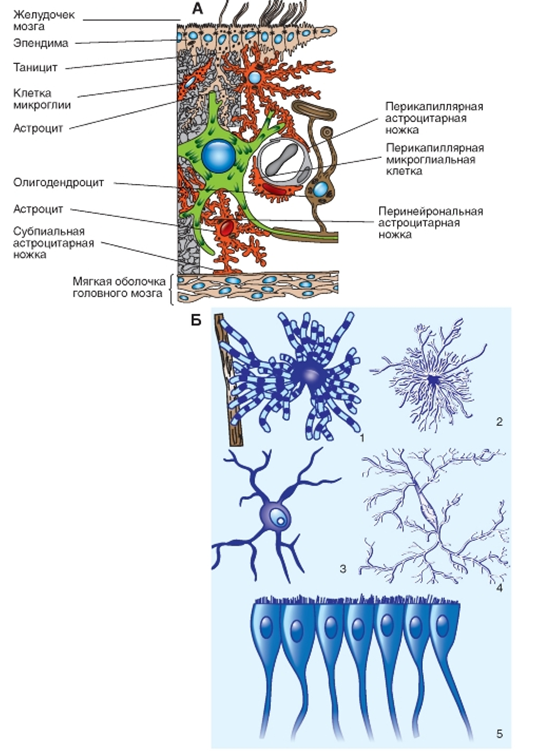 Рис. 6. Ненейронные клетки. А - схематическое представление ненейронных элементов ЦНС: два астроцита, ножки отростков которых заканчиваются на соме и дендритах нейрона, а также контактируют с мягкой мозговой оболочкой и/или капиллярами; олигодендроцит формирует миелиновую оболочку аксонов; клетки микроглии и клетки эпендимы. Б - разные типы клеток нейроглии в ЦНС: 1 - фибриллярный астроцит; 2 - протоплазматический астроцит; 3 - олигодендроцит; 4 - клетки микроглии; 5 - клетки эпендимы
Рис. 6. Ненейронные клетки. А - схематическое представление ненейронных элементов ЦНС: два астроцита, ножки отростков которых заканчиваются на соме и дендритах нейрона, а также контактируют с мягкой мозговой оболочкой и/или капиллярами; олигодендроцит формирует миелиновую оболочку аксонов; клетки микроглии и клетки эпендимы. Б - разные типы клеток нейроглии в ЦНС: 1 - фибриллярный астроцит; 2 - протоплазматический астроцит; 3 - олигодендроцит; 4 - клетки микроглии; 5 - клетки эпендимы Астроциты (звездчатая форма) регулируют микросреду вокруг нейронов ЦНС, их отростками окружены группы синаптических окончаний, которые в результате изолированы от соседних синапсов. Особые отростки - «ножки» астроцитов образуют контакты с капиллярами и с соединительной тканью на поверхности ЦНС - с мягкой мозговой оболочкой (рис. 6). Ножки ограничивают свободную диффузию веществ в ЦНС. Астроциты могут активно поглощать К+и нейромедиаторные вещества, затем метаболизируя их. Таким образом, астроциты играют буферную роль, перекрывая прямой доступ для ионов и нейромедиаторов во внеклеточную среду вокруг нейронов. В цитоплазме астроцитов находятся глиальные филаменты, выполняющие в ткани ЦНС механическую опорную функцию. В случае повреждения отростки астроцитов, содержащие глиальные филаменты, подвергаются гипертрофии и формируют глиальный «рубец».
| |
Миелин увеличивает скорость проведения потенциалов действия благодаря тому, что ионные токи во время потенциала действия входят и выходят только в перехватах Ранвье (областях перерыва между соседними миелинизирующими клетками). Таким образом, потенциал действия «перескакивает» от перехвата к перехвату - так называемое сальтаторное проведение.
Кроме того, в состав нейроглии входят клетки-сателлиты, инкапсулирующие нейроны ганглиев спинальных и черепных нервов, регулируя микросреду вокруг этих нейронов наподобие того, как это делают астроциты.
Еще один вид клеток - микроглия, или латентные фагоциты. В случае повреждения клеток ЦНС микроглия способствует удалению продуктов клеточного распада. В этом процессе участвуют другие клетки нейроглии, а также фагоциты, проникающие в ЦНС из кровотока.
Ткань ЦНС отделена от ЦСЖ, заполняющей желудочки мозга, эпителием, сформированным эпендимными клетками (рис. 6). Эпендима обеспечивает диффузию многих веществ между внеклеточным пространством мозга и ЦСЖ. Специализированные эпендимные клетки сосудистых сплетений в системе желудочков секретируют значительную долю ЦСЖ.
Тест
1. Выберите наиболее точный ответ. В ЦНС присутствуют клетки, обеспечивающих функцию анализа информации и передачи ее эффекторным клеткам
a) нейроны и глиальные клетки
b) различные типы нейронов
2. Верно ли, что миелиновая оболочка нервных волокон обеспечивает более высокую скорость проведения возбуждения
a) да
b) нет
3. Глиальные клетки выполняют
a) разнообразные функции
b) только функцию опоры и питания для нейронных сетей
- Физиология синапсов. Типы. Строение. Механизмы передачи возбуждения
Электрическое взаимодействие клеток: электрический синапс
На рисунке 7А схематично представлена клетка с основными известными типами контактных структур.
1. Плотный контакт (tight junction) характеризуется слиянием внешних листков мембран соседних клеток с образованием одиночной электронно-плотной полосы вдоль контакта и представляет собой сеть ветвящихся тонких гребней
2. Непосредственно к плотному контакту примыкает зона слипания, или промежуточный контакт (intermediate junction), заполненный гомогенным материалом низкой электронной плотности.
3. Десмосома (desmosome) - локальное дискообразное электронно-плотное образование, располагающееся параллельно внутреннему листку каждой клеточной мембраны. От каждой десмосомы внутрь цитоплазмы расходятся пучки фибрилл.
4. Щелевой контакт (gap junction) - область тесного сближения мембран (20-4-А) двух соседних клеток. Представляет интерес для физиологии с позиций межклеточного взаимодействия.
Щелевой контакт, или gap junction, - тип контактов между клетками как электровозбудимых, так и электроневозбудимых тканей. В бислое мембран обеих клеток, образующих щелевой контакт, белковая часть представлена цилиндрическими структурами, распространяющимися по всей ширине щели и пронизывающими насквозь оба бислоя. Эти, заполненные водой каналы, служат основой для осуществления физиологических функций щелевого контакта (рис. 7Б ).
Щелевой контакт состоит из субъединиц - коннексонов - с расстоянием между ними 80-100 А, представленных 6 коннексинами полипептидной природы, создающих водный канал между цитоплазмой двух соседних клеток. Одиночные коннексины каждого коннексона могут изгибаться относительно друг друга, открывая или закрывая центральный канал коннексона (рис.7В).
| |
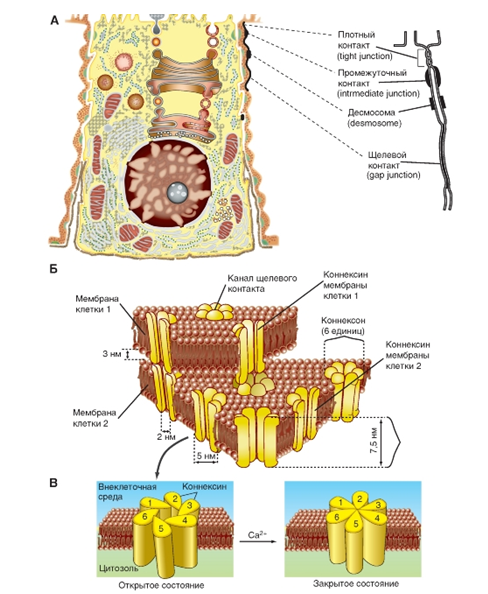 Рис. 7. Межклеточные контакты и структура щелевого контакта.
Рис. 7. Межклеточные контакты и структура щелевого контакта. А - типы межклеточных контактов. Б - модель структуры щелевого контакта, включающая липидный бислой двух соседних клеток, содержащий коннексоны, каждый из которых построен из шести коннексинов. В - в увеличенном виде представлено открытое и закрытое состояние канала коннексона
Структура коннексонов (рис. 8)
Коннексоны позволяют осуществлять обмен ионами и водорастворимыми молекулами с молекулярной массой до 1200-1500 Да между клетками: возможность метаболической кооперации между клетками, например, АМФ, АДФ или АТФ, цАМФ(вторичный мессенджер) могут проходить через щелевой контакт. Эти каналы служат основой и для электротонического взаимодействия между клетками. Центральный канал коннексона может закрываться в результате увеличения концентрации внутриклеточного Са2+ или Н+ в одной из клеток, а также в ответ на деполяризацию одной или обеих клеток.
В целом, щелевые контакты обладают следующими свойствами и функциями:
- ответственны за эффективную диффузионную связь между клетками;
- пропускают гидрофильные соединения, не способные проходить через другие участки поверхностных мембран;
- диффузионные каналы между двумя клетками изолированы от остальной межклеточной среды;
- проницаемость контактирующих мембран в области щелевых контактов резко падает при увеличении концентрации свободного внутриклеточного кальция, закисления или деполяризации любой из клеток;
- диффузионные каналы из клетки в клетку способны формироваться в течение десятков секунд или минут при тесном сближении мембран двух соседних клеток.
| |
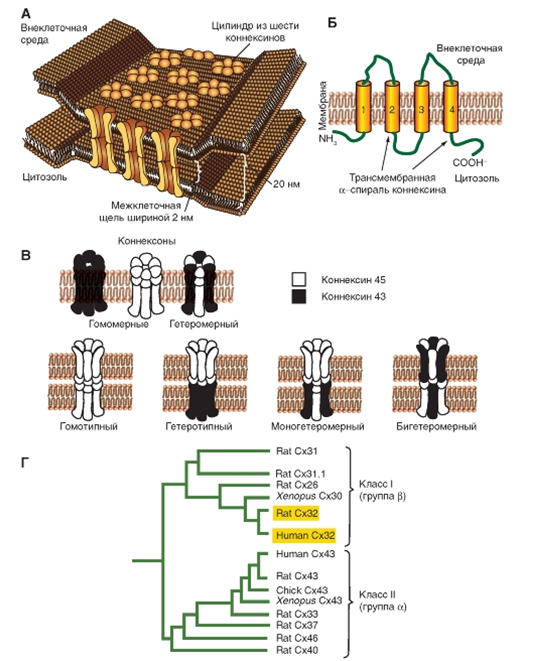 Рис. 8. Коннексоны и их молекулярная организация
Рис. 8. Коннексоны и их молекулярная организация Роль щелевого контакта в проведении возбуждения в ткани
Мембрана электровозбудимых клеток потенциалуправляема, т.е. работа ионных каналов управляется потенциалом, а изменение потенциала зависит от тока, поступающего, например, через щелевой контакт от соседних клеток. При достижении потенциала порога электровозбудимые клетки генерируют потенциалы действия, распространяющиеся, например у нейронов, по отросткам к другим нервным клеткам или эффекторным органам, у мышечных клеток запускают сокращение, а у возбудимых клеток желез управляют секрецией.
Таким образом, функционирование возбудимых тканей связано с распределением потенциала и распространением тока в них.
Транспорт веществ через щелевой контакт
Одно из важнейших свойств щелевого контакта как транспортной системы состоит в том, что при его наличии между соседними клетками может осуществляться интенсивный диффузионный обмен гидрофильными соединениями с молекулярной массой до 1500 Да, минуя наружную среду. Через коннексоны щелевого контакта проходят неорганические ионы (Na+, K+, Cl-, I-, SO42-), сахара (мальтоза, мальтотриоза, мальтотетроза, сахароза), аминокислоты (аспарагиновая, глутаминовая, гексоглицин), нуклеотиды (гипоксантин, аденин, уридин, тиогуанин). Многие из перечисленных веществ не способны диффундировать через наружную мембрану клеток.
Электрический синапс
Механизм передачи сигнала через электрический синапс аналогичен механизму распространения потенциала действия по нервному волокну. В нервном волокне потенциал действия возникает за счет разности потенциалов между возбужденной и невозбужденной областями. Это вызывает открытие Na+-каналов и генерацию импульса заново на каждом последующем участке волокна. В электрическом синапсе в результате разности потенциалов между возбужденным и невозбужденным участками возникают локальные токи, и потенциал из возбужденной терминали аксона распространяется в постсинаптическую клетку, пройдя через ее мембрану и замыкаясь снаружи. Это приводит к открытию Na+-каналов в мембране постсинаптической клетки и возникновению там потенциала действия. Такой механизм работы требует низкого сопротивления пре- и постсинаптической мембран, что обеспечивается наличием коннексонов.
| |
- Быстродействие, что позволяет обеспечивать быстрые реакции организма.
- Синхронизация работы нейронов.
- Возникновение импульсных разрядов в группе электрически связанных клеток.
- Выпрямление сигнала, что обеспечивает его передачу только в одном направлении.
В сердце электрическая связь показана для клеток всех его отделов, в том числе между клетками с разной дифференцировкой, например между волокнами Пуркинье и клетками рабочего миокарда. Поскольку щелевые контакты между клетками обладают низким сопротивлением относительно мембраны контактирующих клеток, возбужденная клетка может передавать сигнал невозбужденной, в результате чего эта последняя возбуждается.
| |
Лиганд-рецепторное взаимодействие. Лиганд - это молекула, которая может взаимодействовать с участком связывания (местами связывания) молекулярного рецептора - специфическим участком молекулы белковой природы, встроенной, например, в мембрану клетки или находящейся в цитоплазме.
| |
Взаимодействие лиганда с участком связывания молекулярного рецептора может быть строго специфичным, и, следовательно, участок связывания может связывать только один тип лиганда.
Хотя некоторые участки связывания имеют химическую специфичность, которая позволяет им связывать только один тип лиганда, другие участки менее специфичны и, таким образом, способны связывать ряд родственных лигандов, поскольку фрагмент каждого такого лиганда может быть комплементарен форме участка связывания.
Сила связывания лиганда с участком связывания белка известна как его аффинность или сродство. Мерой сродства участка связывания к лиганду служит концентрация лиганда, необходимая для обеспечения 50% насыщения; чем более низкая концентрация лиганда необходима для связывания его с половиной центров связывания, тем больше афинность участка связывания к лиганду.
| |
| |
Агонист - это лиганд, который взаимодействует с участком связывания и изменяет состояние молекулярного рецептора подобно трансмиттеру.
Наряду с агонистами существуют химические соединения, которые хотя и взаимодействуют с участком связывания на макромолекуле рецептора, но не могут открыть ионный канал. Занимая участок связывания, они препятствуют рецепции естественного трансмиттера и возникновению его эффекта, поскольку действуют на ту же самую макромолекулу рецептора, что и агонист. Такие вещества называют антагонистами.
| |
Так как в формировании потенциала покоя мембраны участвует большое число ионов, равновесие может нарушаться посредством изменений проводимости различных ионов. Так, например, при дополнительном выходящем токе ионов K+ или при входящем токе ионов Cl-может происходить гиперполяризация, которая является противоположностью возбуждения, вызывающей торможение нейрона.
| |
Химические процессы в синапсе открывают широкие возможности для фармакологической регуляции и оказываются предметом многочисленных исследований с целью поиска эндогенных соединений, способных модифицировать в заданных направлениях синаптическую передачу.
И действительно, многие медикаменты реализуют свое фармакологическое действие путем влияния на синаптическое проведение. Это относится не только к психотропным и наркотическим веществам. Многие другие, например, (гипотензивные) средства понижающие артериальное давление, также действуют опосредованно через синапсы. Кроме того, многие яды растительного и животного происхождения направленно действуют на химический синапс.
Почти все синаптические терминали освобождают не один только трансмиттер, но одновременно с ним целый ряд биологически активных соединений. Примерами таких сопутствующих соединений служат АТФ, ГТФ, окситоцин, вещество Р, энкефалин и др. Их называют котрансмиттеры.
Многие трансмиттеры, например, глицин и ГАМК или глицин и глутамат могут присутствовать в везикулах одновременно.
Котрансмиттеры могут модулировать синаптические процессы. Накапливаясь в везикулах, они обладают собственной кинетикой освобождения, например освобождение только при значительной величине пресинаптического потенциала.
| |
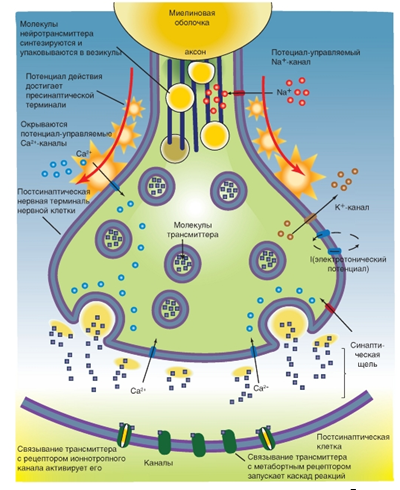 Рис. 9. Химический синапс и принцип его работы
Рис. 9. Химический синапс и принцип его работы Механизм высвобождения трансмиттера. Во время деполяризации в нервном окончании возникает не только входящий ток Na+. Мембрана окончания аксона имеет и потенциал-управляемые Ca2+-каналы, через которые во время деполяризации ионы Ca2+ проникают в синаптическое окончание. Одновременно ионы Ca2+ дополнительно выходят из эндоплазматического ретикулума. В синаптическом окончании в зоне пресинаптической мембраны расположено большое число синаптических пузырьков (везикул) с медиатором (трансмиттером).
Преобразование трансмиттера. Трансмиттер может быть либо быстро химически расщеплен на неактивные компоненты, либо удален из синаптической щели путем высокоселективного обратного захвата в пресинаптическое окончание. В центральной нервной системе клетки глии могут также захватывать трансмиттер.
Пресинаптические окончания благодаря описанному механизму обратного захвата (реаптейка) захватывают и возвращают либо фрагменты трансмиттера (например, холин), либо всю молекулу трансмиттера (например, серотонин). Для этого в пресинаптической мембране находятся специфические протеины - транспортеры.
Во многих синапсах рецепторы для трансмиттера есть не только в постсинаптической мембране, но также и в пресинаптической. Это так называемые ауторецепторы.
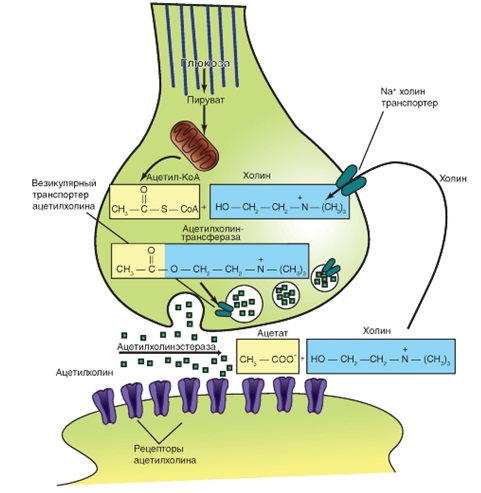 Рис. 10. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
Рис. 10. Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина Типы химических синапсов.
У части синапсов ионный канал, который при действии лиганда, т.е. благодаря связыванию с молекулой трансмиттера, открывается (каналы, управляемые лигандами), называется ионотропным рецептором (рис. 11 А).
| |
Примером и ионотропного, и метаботропного синапса служит холинергический синапс, у которого трансмиттером служит АЦХ. У холинергических синапсов два типа рецепторов: АЦХ-рецепторы, чувствительные дополнительно к никотину (никотиновый АЦХ-рецептор), и АЦХ-рецепторы, чувствительные дополнительно к мускарину (мускариновый АЦХ-рецептор). Первый является ионотропным, второй - метаботропным.
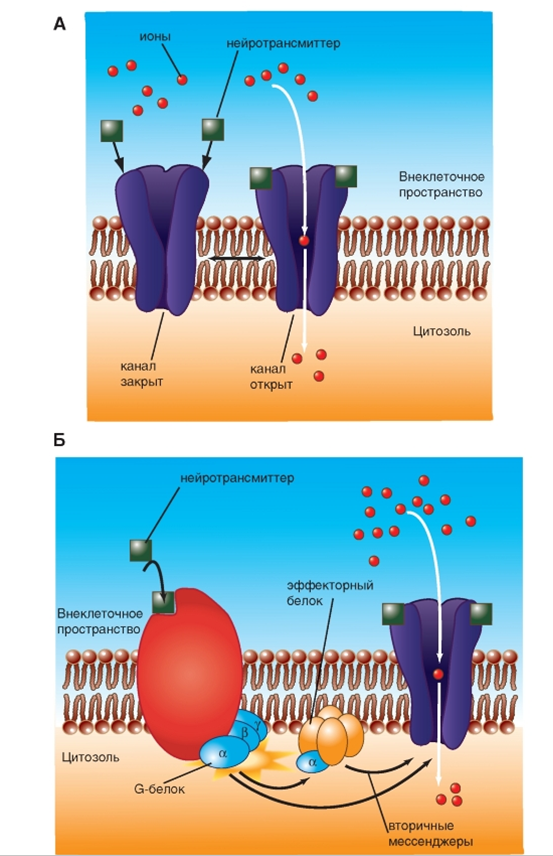 Рис. 11. Ионотропные и метаботропные синапсы
Рис. 11. Ионотропные и метаботропные синапсы 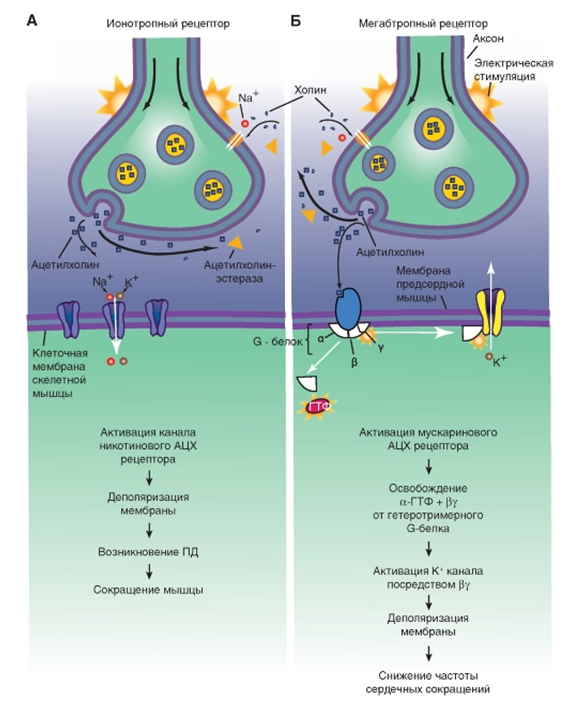 Рис. 12. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б)
Рис. 12. Механизмы работы ионотропного синапса (А) и метаботропного синапса (Б) Тест
1. В ЦНС человека ключевую роль в передаче информации между клектами играют
a) электрические и химические синапсы
b) химические синапсы
2. Основу электрического синапса у человека играют
a) щелевые контакты
b) десмосомы и щелевые контакты
3. В структуре химического синапса переача сигнала осуществляется посредством
a) нейромедиатора
b) коннексинов
4. Химические синапсы ЦНС представлены следующими типами
a) метаботропными и ионотропными
b) только ионотропными
5. В основе работы метаботропного синапса лежит
a) механизм вторичных мессенджеров
b) изменение проницаемости ионных каналов
- Нейромедиаторы. Нейромодуляторы
Ацетилхолин. Является медиатором для всех двигательных нейронов, многих интернейронов ЦНС, а также нейронов парасимпатической нервной системы. Существует 2 типа рецепторов ацетилхолина: АЦХ-рецепторы, чувствительные к никотину: никотиновый АЦХ-рецептор и, соответственно, никотиновый АЦХ (нАЦХ) синапс, и АЦХ-рецепторы, чувствительные к мускарину - мускариновый АЦХ-рецептор и, соответственно, мускариновый (мАЦХ) АЦХ синапс. АЦХ синапс с никотиновым рецептором являются ионотропными, а АЦХ синапсы с мускариновым рецептором являются метаботропными.
Глутамат в синапсах выполняет функцию трансмиттера и ко-трансмиттера (может присутствовать одновременно с другими трансмиттерами, например с глицином). Глутамат в мозге является важнейшим трансмиттером для возбуждающих синапсов. Синапсы, использующие в качестве трансмиттера глутамат, находятся приблизительно на 50% нейронов центральной нервной системы. Такие синапсы образуют важнейшие возбуждающие входы систем мозга к коре большого мозга. Они принимают участие в процессах обучения. Глутамат прямо открывает неспецифический ионный канал для катионов.
Глицин служит трансмиттером тормозных синапсов и действует как нейромодулятор. Глицин предназначен для выполнения специфических ингибиторных задач. Большинство клеток Реншоу, через которые тормозятся α-мотонейроны спинного мозга, выбрасывают глицин. Глициновые рецепторы открывают каналы для ионов Cl-, что способствует возникновению так называемых тормозных постсинаптических потенциалов (тормозных потенциалов синаптической пластинки - ТПСП). Синаптическое действие заканчивается с помощью механизма обратного захвата глицина.
| |
γ-Аминомасляную кислоту (ГАМК) выбрасывают в качестве трансмиттера многие тормозные интернейроны, имеющиеся практически во всех отделах центральной нервной системы, и аксоны клеток Пуркинье мозжечка. ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей мере два различных постсинаптических механизма, которые запускаются благодаря различным рецепторам (ГАМКА и ГАМКС с одной стороны; ГАМКВ с другой стороны).
Серотонин (5-hydroxytryptamin) - биогенный амин, широко распространенный в ЦНС. В мозге его особенно много в области тектума. Оттуда наблюдается много проекций в лимбическую систему, к таламусу и гипоталамусу, в передний мозг, в мозжечок и в спинной мозг. По этим путям, очевидно, осуществляется контроль многих нейрональных функций. На постсинаптической мембране были обнаружены различные серотониновые рецепторы, которые большей частью, благодаря вторичным мессенджерам, открывают каналы для ионов К+ и Са2+. Действие трансмиттера заканчивается включением механизма обратного захвата пресинаптической областью.
Гистамин - моноамин, выступающий в качестве трансмиттера. Особенно важную роль он играет как модулятор в мозге грудных детей. Гистаминергические нейроны у взрослых находятся в заднем гипоталамусе и связаны со многими участками мозга, где они оказывают влияние на состояние бодрствования, мышечную активность, прием пищи, сексуальные отношения и обменные процессы в мозге. Из-за участия этих нейронов в регуляции процессов сна и бодрствования многие антигистаминные препараты вызывают состояние сонливости.
| |
Дофаминергические пути, которые исходят от area ventalis tegmentalis и проецируются к nucleus accumbeus,играют большую роль в патогенезе эпилепсии.
Существуют различные дофаминовые рецепторы, действующие через вторичные мессенджеры. Их постсинаптическое действие может быть тормозным или возбуждающим. Дофамин очень быстро захватывается обратно из синаптической щели в пресинаптическое окончание. Там он претерпевает превращения благодаря моноаминоксидазе. Вне нейрона, он подвергается воздействию катехол-О-метилтрансферазы.
Норадреналин катехоламин, выполняющий функцию трансмиттера в ЦНС и синтезирующийся прежде всего нейронами locus coeruleus. Это ядро состоит не более чем из 1000 клеток, аксоны которых так многократно разветвляются, что можно найти адренергические окончания во многих областях ЦНС. Они оказывают модулирующее влияние, например, на процессы созревания, обучения, переработку информации в мозгу, регуляцию сна и на эндогенное торможение боли. В периферической нервной системе норадреналин служит важным трансмиттером симпатических постганглионарных окончаний, например, на сердце и на гладких мышцах сосудов. В некоторых центральных синапсах как трансмиттер рассматривают адреналин.
Для катехоламинов существует четыре главных типа рецепторов: α1, α2, β1 и β2. Они отличаются по реакции на различные агонисты или антагонисты, а также по постсинаптическим эффектам. Рецепторы α1 управляют Са2+-каналами при помощи вторичного мессенджера IP3 и при активации повышают внутриклеточную концентрацию ионов Са2+. Активация рецепторов ведет к уменьшению концентрации вторичного мессенджера цАМФ, вызывая различные эффекты. Активация рецепторов β, например, на клетках Пуркинье мозжечка, повышает при помощи вторичного мессенджера цАМФ проводимость мембраны для ионов К+ и, таким образом, образует ТПСП. Обратный захват и разрушение похожи на таковые для дофамина.
Олигопептиды, которые действуют как трансмиттеры или нейромодуляторы - это приемущественно короткие полипептиды, состоящие из 2-30 аминокислотных остатков. Они функционируют как трансмиттеры или ко-трансмиттеры либо модулируют синаптические процессы. Так энкефалин, эндорфин и динорфин служат трансмиттерами в тех синапсах, в которых опиоидные вещества также оказывают действие. Опиоиды (например, морфин) являются сильными анальгетиками. Вышеназванные опиоидные пептиды тормозят распространение боли в спинном мозге. Кроме того, они играют большую роль в лимбической, автономной и моторной системах.
Другие нейропептиды: вещество Р, ангиотензин II, соматостатин, вазоактивный интестинальный полипептид (VIP), нейропептид Y и многие другие. Большинство названных веществ были открыты в качестве гормонов прежде, чем была определена их роль в синаптическом переносе.
ГАМК как трансмиттер
Многие тормозные интернейроны, присутствующие практически во всех отделах ЦНС, и аксоны клеток Пуркинье мозжечка выбрасывают в качестве трансмиттера γ-аминомасляную кислоту (ГАМК, GABA: γ-Aminobutyric acid). ГАМК оказывает тормозное действие на постсинаптические структуры. Существует по меньшей мере два различных постсинаптических механизма, запускающихся благодаря различным рецепторам (ГАМКА, ГАМКС с одной стороны; ГАМКВ с другой стороны).
ГАМКА-и ГАМКС-рецепторы непосредственно открывают каналы для ионов Cl-, что приводит к появлению входящего Cl-тока и тем самым к гиперполяризации в виде ТПСП. Барбитураты (препараты, применяемые для наркоза), стероидные анестетики и транквилизаторы (успокаивающие средства) из класса бензодиазепинов (например, диазепам) усиливают ингибиторное действие ГАМКА-рецепторов. Механизм их действия заключается в связывании с белком ионного канала, который уже взаимодействует с ГАМК, и таким образом в усилении действия ГАМК.
Канал построен из нескольких субъединиц. Одна из этих субъединиц связывает ГАМК, другая может дополнительно связать бензодиазепин, третья - связать барбитурат и еще одна - связать стероид. В настоящее время неясно, какие природные вещества (например, нейромодуляторы) оказывают действие на так называемые бензодиазепиновые или барбитуратные рецепторы. Названные субъединицы могут обладать различными химическими свойствами, определяемыми их химической структурой, что обусловливает различные фармакологические эффекты. БикукуллинÄ представляет собой конкурентный блокатор для ГАМКА-, но не для ГАМКС-рецептора. Яд пикротоксин, вызывающий судороги, является неконкурентным блокатором, закрывающим канал для ионов Cl-.
ГАМКВ открывают каналы для ионов К+ посредством G-белка. Агонистом служит баклофен. Пресинаптические ГАМКВ-рецепторы закрывают каналы Са2+и тормозят освобождение трансмиттера. Действие ГАМК прекращается путем обратного захвата ГАМК пресинаптическим окончанием и клетками глии.
| |
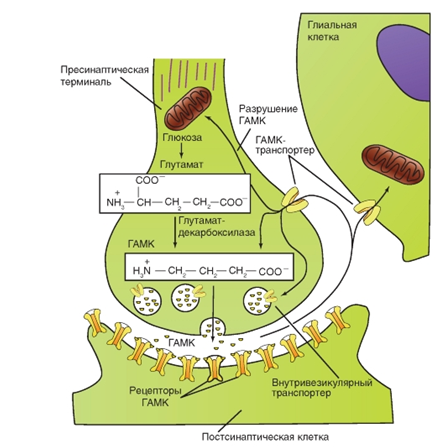 Рис. 13. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный захват
Рис. 13. γ-Аминомасляная кислота как трансмиттер в синапсе: расщепление, удаление и обратный захват Глицин как трансмиттер
Аминокислота глицин выполняет функцию трансмиттера в тормозных синапсах и действует как нейромодулятор. Глицин выполняет специфические ингибиторные задачи. Большинство клеток Реншоу, через которые осуществляется торможение α-мотонейронов спинного мозга, высвобождают глицин. Стрихнин является конкурентным антагонистом, и его применение приводит к судорогам из-за отсутствия торможения α-мотонейронов. Рецепторы глицина открывают С1--каналы, что способствует возникновению ТПСП. Синаптическое действие заканчивается с помощью механизма обратного захвата глицина.
В ЦНС глицин играет также и роль нейромодулятора, например в гиппокампе NMDA-рецепторы, описанные ранее, испытывают модулирующее влияние со стороны глицина.
Важно заметить, что хотя синапсы называются по наименованию их главного трансмиттера (например, холинергический).
NO как трансмиттер
Оксид азота (NO) - это паракринный медиатор, выделяемый эндотелиальными клетками и некоторыми нейронами. Вследствие того, что NO быстро окисляется, его биологическая жизнь длится всего несколько секунд. По этой причине NO влияет только на клетки, находящиеся вблизи от места своего выделения. NO стимулирует растворимую гуанилатциклазу в клетках-мишенях и тем самым повышает в клетке мишени внутриклеточную концентрацию цГМФ. Это, в свою очередь, стимулирует цГМФ-зависимую протеинкиназу.
Оксид азота легко распространяется путем диффузии и может принимать на себя функции трансмиттера. При этом, например, пресинаптические процессы могут регулироваться с помощью обратной связи. Возможно, этот вид переноса информации играет большую роль в ЦНС. Механизмы прекращения действия подобных трансмиттеров не позволяют включить их в классическую синаптическую схему.
Тест
1. Что из перечисленного не является нейротрансмиттером?
a) ацетилхолин, норадреналин, ГАМК, серотонин
b) холинэстераза, катехол-О-метилтрасфераза
2. Верно ли следующее утверждение. Оксид азота не является нейротрансмиттером т.к. это вещество очень быстро окисляется
a) да, верно
b) нет, не верно
3. Верно ли утверждение, что каждый нейротрансмиттер может связываться только с ионотропным или только с метаботропным рецептором?
a) да, верно
b) нет, не верно
5. Постсинаптические потенциалы
Схема распространения информации по нейрону (рис.14)
В зоне синапса локальный возбуждающий постсинаптический потенциал (ВПСП) распространяется пассивно электротонически по всей постсинаптической мембране клетки. Это распространение не подчиняется закону «все или ничего». Если большое число возбуждающих синапсов возбуждаются одновременно или почти одновременно, то возникает явление суммации, проявляющееся в виде возникновения ВПСП существенно большей амплитуды, что может деполяризовать мембрану всей постсинаптической клетки. Если величина этой деполяризации достигает в области постсинаптической мембраны порогового значения (10 мВ или больше), то на аксонном холмике нервной клетки молниеносно открываются потенциалуправляемые Na+-каналы, и клетка генерирует потенциал действия, проводящийся вдоль ее аксона. От начала ВПСП до образования потенциала действия проходит еще около 0,3 мс.
Пороговый стимул - самый слабый стимул, надежно различаемый сенсорным рецептором. Для этого стимул должен вызывать рецепторный потенциал такой амплитуды, которая достаточна для активации хотя бы одного первичного афферентного волокна. Более слабые стимулы могут вызвать подпороговый рецепторный потенциал, однако они не приведут к возбуждению центральных сенсорных нейронов и, следовательно, не будут восприняты. Кроме того, количество возбужденных первичных афферентных нейронов, необходимое для сенсорного восприятия, зависит от пространственной и временной суммации в сенсорных путях (рис. 14, Г).
Взаимодействуя с рецептором, молекулы АЦХ открывают неспецифические ионные каналы в постсинаптической мембране клетки так, что повышается их способность к проводимости одновалентных катионов. Работа каналов ведет к базовому входящему току положительных ионов, и, следовательно, к деполяризации постсинаптической мембраны, которая применительно к синапсам называется возбуждающим постсинаптическим потенциалом.
| |
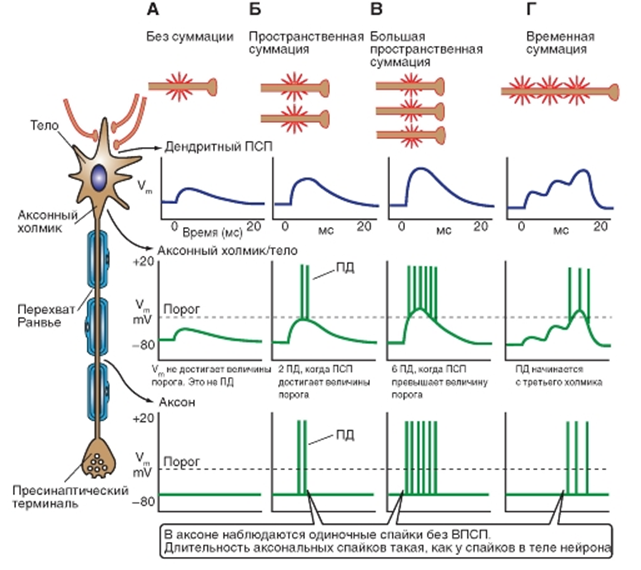 Рис. 14. Информация течет от дендритов к соме, к аксону, к синапсу. На рисунке представлены типы потенциалов в разных местах нейрона в зависимости от пространственной и временной суммации
Рис. 14. Информация течет от дендритов к соме, к аксону, к синапсу. На рисунке представлены типы потенциалов в разных местах нейрона в зависимости от пространственной и временной суммации Возбуждающий постсинаптический потенциал (ВПСП)
Рассмотрим этот вопрос на примере АЦХ. Взаимодействуя с рецептором, молекулы АЦХ открывают неспецифические ионные каналы в постсинаптической мембране клетки так, что повышается их способность к проводимости для одновалентных катионов. Существует сильная движущая сила для натрия, и ионы Na+устремляются внутрь клетки и деполяризуют ее мембрану. С другой стороны, канал проходим и для ионов K+, для которых сохраняется незначительный электрохимический градиент, направленный из внутриклеточной области к внеклеточной среде. Через постсинаптическую мембрану одновременно проходят и ионы K+, тем самым слегка противодействуя деполяризации, обусловленной входящим током ионов Na+. Работа описанных каналов ведет к базовому входящему току положительных ионов и, следовательно, к деполяризации постсинаптической мембраны, которая называется применительно к синапсам ВПСП. На концевой пластинке нервно-мышечного синапса ВПСП называют также потенциалом концевой пластинки. Так как участвующие ионные токи зависят от разности их потенциала равновесия и потенциала мембраны, то при уменьшенном потенциале покоя мембраны ток ионов Na+ становится всегда меньше, а ток ионов К+- больше. Именно поэтому амплитуда ВПСП уменьшается.
| |
Прежде всего, в зоне синапса образовавшийся локально ВПСП распространяется пассивно электротонически по всей постсинаптической мембране клетки. Этот процесс не подчиняется закону «все или ничего». Если большое число возбуждающих синапсов активируются одновременно или почти одновременно, то возникает явление так называемой суммации, которое реализуется в виде возникновения ВПСП существенно большей амплитуды, что может деполяризовать мембрану всей постсинаптической клетки. Если величина этой деполяризации достигает в области постсинаптической мембраны порогового значения (10 мВ или больше), то на аксонном холмике нервной клетки молниеносно открываются потенциал-управляемые Na+-каналы, и клетка генерирует потенциал действия, который проводится вдоль аксона. В случае моторной концевой пластинки это приводит к мышечному сокращению. От начала ВПСП до образования потенциала действия проходит еще около 0,3 мс, так что при обильном освобождении трансмиттера постсинаптический потенциал может появиться уже через 0,5-0,6 мс после пришедшего в пресинаптическую область потенциала действия. В общих чертах, время «синаптической задержки», подразумевающее необходимое время между возникновением пре- и постсинаптического потенциала действия, всегда зависит от типа синапса.
| |
Решающими для ионного тока в мембране оказываются величина ее потенциала и количество открытых ионных каналов. Определяющим оказывается тип белка канала, на который действует трансмиттер. Так на одних синапсах находятся каналы для K+, тогда как на других есть каналы для Cl-. Последние распространены много чаще. Рассмотрим в виде примера рецептор метаботропного синапса, повышающий при связывании с трансмиттером проводимость для ионов К+. При нормальной величине потенциала мембраны это приводит к дальнейшему выходящему току ионов К+ и гиперполяризации потенциала мембраны. Возникает так называемый ТПСП. Принципиально похожая ситуация складывается, если гиперполяризующий мембрану ток связан с ионами Cl-. Cl- течет в клетку и гиперполяризует ее. Подобная картина характерна для очень многих клеток.
| |
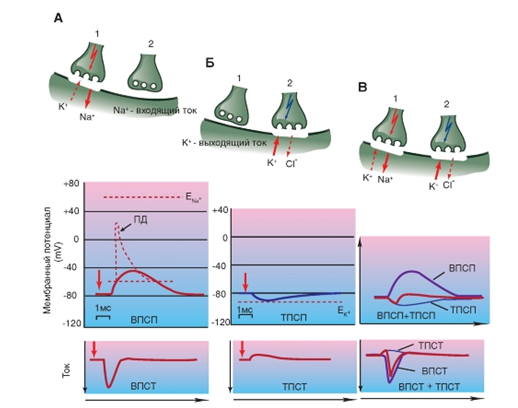 Рис. 15. Потенциалы и токи, возникающие на постсинаптической мембране.
Рис. 15. Потенциалы и токи, возникающие на постсинаптической мембране. А - процессы освобождения трансмиттера и формирование возбуждающего постсинаптического потенциала (ВПСП) после электрического возбуждения (стрелка), поступившего на пресинаптическое окончание, связаны с возникновением возбуждающего постсинаптического тока (ВПСТ) и генерацией вследствие этого ВПСП. Если ВПСП переходит порог, то возникает потенциал действия (ПД), который во время фазы нарастания благодаря потенциалуправляемым Na+каналам деполяризует мембрану в направлении ENa, т.е. к потенциалу равновесия для Na+. Б - процессы освобождения трансмиттера и формирование ТПСП связаны с ингибиторным постсинаптическим током (ТПСТ) и соответствующим ему ТПСП. При этом ток вызывает движение через мембрану ионов К+. Ионы Cl- могут играть роль, если потенциал мембраны позитивнее, чем потенциал равновесия для Cl-, который лежит от -75 до -70 мВ. В - токи мембраны при активации возбуждающих (ВПСТ) и ингибиторных синапсов (ТПСТ) и возникающие в этом случае результирующие постсинаптические потенциалы (ВПСП, ТПСП). При одновременной активации возбуждающих и тормозных синапсов возникающие мембранные токи суммируются, поэтому результирующий постсинаптический потенциал (красный) становится очень маленьким
Взаимное влияние ВПСП и ТПСП
Если на мембране клетки активируются одновременно возбуждающие и тормозные синапсы, то ионный ток уменьшается.
Если соседствующие возбуждающие и тормозные синапсы (одновременно) активируются, то возникающие токи накладываются друг на друга. Результирующий постсинаптический потенциал (по абсолютной величине) меньше, чем только один ВПСП или только один ТПСП.
| |
Тест
1. На постсинаптической мембране могут возникать как возбуждающие, так и тормозные потенциалы, что зависит от
a) типа медиатора и рецепторов, открывающих разные ионные каналы
b) только от типа медиатора
2. Возбуждающий постсинаптический потенциал связан с открытием
a) натриевых и кальциевых ионных каналов
b) хлорных и калиевых ионных каналов
3. Тормозный постсинаптический потенциал связан с открытием
a) натриевых и кальциевых ионных каналов
b) хлорных и калиевых ионных каналов
4. ВПСП – это всегда
a) деполяризация мембраны
b) гиперполяризация мембраны
5. ТПСП – это всегда
a) гиперполяризация мембраны
b) деполяризация мембраны
- Аксональный транспорт
Существуют несколько типов аксонального транспорта. Окруженные мембранами органоиды и митохондрии транспортируются с относительно большой скоростью посредством быстрого аксонального транспорта. Вещества, растворенные в цитоплазме (например, белки), перемещаются с помощью медленного аксонального транспорта. У млекопитающих быстрый аксональный транспорт имеет скорость 400 мм/сутки, а медленный аксональный транспорт - около 1 мм/сутки.
Для аксонального транспорта требуется затрата метаболической энергии и присутствие внутриклеточного Са2+. Элементы цитоскелета (точнее, микротрубочки) создают систему направляющих тяжей, вдоль которых передвигаются окруженные мембранами органоиды.
| |
Высокая, в ряде случаев, потребность в молекулах трансмиттера вызывает, как правило, синтез трансмиттера на месте, то есть непосредственно в пресинаптическом окончании.
| |
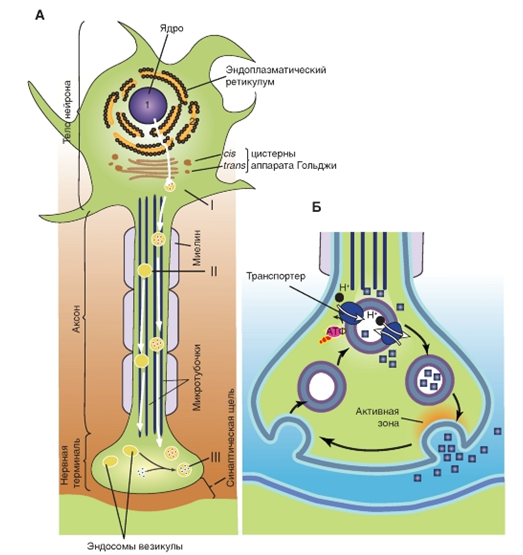 Рис. 16. Синтез и рецикл синаптических везикул и их содержимого.
Рис. 16. Синтез и рецикл синаптических везикул и их содержимого. А - Биосинтез нейротрансмиттеров.
I. Везикулы, предшественники пептидных трансмиттеров и ферменты синтезируются в теле нейрона и высвобождаются из аппарата Гольджи.
II. Везикулы перемещаются по аксону с помощью быстрого аксонального транспорта. Пептидные трансмиттеры уже заключены в некоторые везикулы.
III. Непептидные нейротрансмиттеры синтезируются и транспортируются в везикулы непосредственно в нервных окончаниях
Б - Экзоцитоз (высвобождение визикул с медиатором)
Тест
1. Органоиды перемещаются вдоль аксона путем
а) быстрого аксонального транспорта
б) медленного аксонального транспорта
2. Молекулы растворенных в цитоплазме нейрона веществ перемещаются вдоль аксона путем
а) медленного аксонального транспорта
б) быстрого аксонального транспорта
3. Антероградный аксональный транспорта имеет направление
а) от сомы к аксональным терминалям
б) от нервного окончания к соме
4. Ретроградный аксональный транспорт имеет направление
а) от сомы к аксональным терминалям
б) от нервного окончания к соме
- Гематоэнцефалический барьер. Цереброспинальная жидкость
Общий объем ЦСЖ в желудочках мозга - примерно 35 мл, тогда как подпаутинное пространство содержит около 100 мл. Каждую минуту образуется примерно 0,35 мл ЦСЖ. При такой скорости обновление ЦСЖ происходит приблизительно четыре раза в сутки.
У человека в положении лежа давление ЦСЖ в спинно-мозговом субарахноидальном пространстве достигает 120-180 мм вод.ст. Скорость образования ЦСЖ относительно независима от давления в желудочках и в субарахноидальном пространстве, а также от системного кровяного давления. В то же время скорость обратного всасывания ЦСЖ прямо связана с давлением ЦСЖ.
Внеклеточная жидкость в ЦНС непосредственно сообщается с ЦСЖ. Следовательно, состав ЦСЖ влияет на состав внеклеточной среды вокруг нейронов головного и спинного мозга. Основные компоненты ЦСЖ в поясничной цистерне перечислены в табл. 5-1. Для сравнения приведены концентрации соответствующих веществ в крови. Как показано в данной таблице, содержание К+, глюкозы и белков в ЦСЖ ниже, чем в крови, а содержание Na+ и Cl- - выше. Кроме того, в ЦСЖ практически нет эритроцитов. Благодаря повышенному содержанию Na+ и Cl- обеспечивается изотоничность ЦСЖ и крови, несмотря на то, что в ЦСЖ относительно мало белков.
Таблица. Состав цереброспинальной жидкости и крови

Характеристики лекций
 СГМУ
СГМУСписок файлов
 Spes
Spes
21 июня 2021 в 12:31
Комментарии
Нет комментариев
Стань первым, кто что-нибудь напишет!
Отзывы на другие работы автора

Отзыв
Софисты, Сократ и Платон. Философия Аристотеля (Семинар №5)
отлично. ответ полный, соответствует вопросу. всем рекомендую
Отзыв
Античная философия. Досократический период (Семинар №4)
отлично подошло. есть вся информация в полном объеме.
Отзыв
Античная философия. Эллинистический период (Семинар №6)
вся информация в полном объеме. отлично все есть. помогло!
Отзыв
Немецкая классическая философия (Семинар №10)
отлично! точное совпадение. помогло!!!)))
Отзыв
Новоевропейская философия XVII-XVIII вв. (Семинар №8)
отлично! все задания совпадают. помогло!!!


















