
Эмбриогенез амниот
Тема 5
Эмбриогенез амниот
1. Общая характеристика амниот.
2. Эмбриогенез пресмыкающихся.
3. Эмбриогенез птиц.
4. Эмбриогенез млекопитающих.
5. Эмбриогенез человека и приматов.
Рекомендуемые материалы
6. Типы плацент.
ИСПОЛЬЗОВАННАЯ И РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
1. Антипчук, Ю.П. Гистология с основами эмбриологии / Ю.П. Антипчук. – М.: Просвещение, 1983. – 240 с.
2. Алмазов, И.В., Сутулов Л.С. Атлас по гистологии и эмбриологии / И.В. Алмазов, Л.С. Сутулов. – М.: Медицина, 1978. – 148 с.
3. Гистология / под ред. Ю.И. Афанасьева. – М: Медицина, 1989. – 361 с.
4. Рябов, К.П. Гистология с основами эмбриологии / К.П. Рябов. – Мн.: Высш. шк., 1991. – 289 с .
5. Биологический энциклопкдический словарь / под ред. М.С. Гилярова. – М.: Сов. Энцикл., 1989. – 864 с.
6. Практикум по гистологии, цитологии и эмбриологии / под ред. Н.А. Юриной, А.И. Радостиной. – М.: Высш. шк., 1989. – 154 с.
Хэм А., Кормик Д. Гистология / А. Хэм, Д. Кормик. – М.: Мир, 1983. – 192
1. Общая характеристика амниот
К амниотам относятся хордовые, у которых в процессе развития возникают такие зародышевые оболочки, как амнион и аллантоис. Эти зародышевые оболочки присущи хордовым, живущим на суше, а также вторичноводным животным из классов пресмыкающихся, птиц и млекопитающих. Развитие зародышей у этих животных, а также у человека происходит в полости, образуемой амниотической оболочкой, которая наполнена амниотической, или околоплодной, жидкостью. Таким образом, эмбриональное развитие амниот, как и анамний, осуществляется в водной среде, что является показателем преемственности филогенетического развития всех хордовых. Однако амниотическая жидкость не может обеспечить развивающемуся зародышу оптимальные условия для дыхания, выделения и питания. Эти функции осуществляет аллантоис, в котором развивается густая сеть кровеносных сосудов, благодаря которым осуществляется газообмен зародыша с окружающей средой и выделение в полость аллантоиса конечных продуктов обмена веществ, как это имеет место у пресмыкающихся, птиц и яйцекладущих млекопитающих. Через аллантоис зародыш других млекопитающих контактирует с телом матери, получая от нее все необходимое для развития.
Эмбриогенез амниот мы рассматриваем на примере развития позвоночных животных трех классов: пресмыкающихся, птиц и млекопитающих. Изучение особенностей развития амниот начинаем с изучения эмбриогенеза пресмыкающихся, поскольку от них в процессе эволюционного развития произошли птицы и млекопитающие. Это нашло отражение в общности ряда стадий эмбрионального развития этих позвоночных животных, а также человека.
2. Эмбриогенез пресмыкающихся
Дробление яйцеклетки у пресмыкающихся неполное, дискоидальное. Так как оплодотворение яйцеклетки происходит в верхней трети яйцевода, то дробление начинается во время движения яйцеклетки по яйцеводу и при откладывании яйца зародыш находится либо на стадии дискобластулы, либо ранней гаструлы. У ряда пресмыкающихся, в связи с длительным нахождением яйца в яйцеводе происходит формирование нового организма, который рождается, или выходит из яйца сразу же после его откладывания (живородящая ящерица, гадюки). Дискоидальное дробление приводит к возникновению дискобластулы. Крыша бластулы состоит из мелких клеток бластодермы, совокупность которых называется бластодиском, а дно бластулы образовано нераздробленной массой желтка. Часть желтка под зародышевым диском резорбируется и образуется подзародышевая полость в виде щели между внешним и внутренним слоем бластодиска.
Первая фаза гаструляции осуществляется путем деляминации, к которой иногда присоединяется иммиграция. При этом глубокие слои клеток бластодиска разрыхляются и отщепляются в подзародышевую полость, образуя внутренний зародышевый слой клеток, или энтодерму, а остальные клетки бластодиска развиваются в эктодерму. Процесс образования энтодермы у амниот сместился на более ранние сроки эмбрионального развития и происходит уже в процессе дробления.
После деляминации клеточного материала бластодиска и образования эктодермы и энтодермы у пресмыкающихся возникает двуслойный зародышевый щиток, или первичная пластинка (рисунок 2.1).
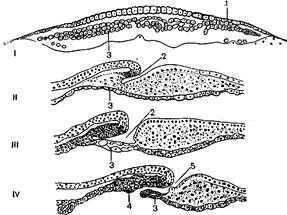
I – первая фаза гаструляцни; II–IV– вторая фаза гаструляции; 1 – первичная пластинка; 2 – гастроцель; 3 – энтодерма; 4 – клеточный материал хорды;
5 – нервно-кишечный канал
Рисунок 2.1 – Гаструляция у пресмыкающихся
Первичная пластинка прогибается, затем впячивается и образует гастральную полость, которая слепым концом направлена вперед, в промежуток между наружным и внутренним зародышевыми листками. Отверстие, через которое впячивается клеточный материал первичной пластинки эктодермального происхождения, является бластопором, а полость впячивания – полостью первичной кишки. Однако на первых порах полость первичной кишки образована клетками наружного зародышевого листка, или эктодермы. Через дорсальную и боковые губы бластопора инвагинирует клеточный материал будущей хорды и мезодермы.
В процессе инвагинации первичной пластинки дно гастральной полости достигает энтодермы. Стенка дна гастральной полости постепенно истончается и в дне гастроцеля, в слое клеток энтодермы возникают отверстия. Эти отверстия увеличиваются, и клеточная перегородка между энтодермой и гастральным впячиванием исчезает. Так возникает единая полость первичной кишки. Клетки энтодермы, размножаясь, вновь соединяются в единый пласт под клеточным материалом хорды и мезодермы. У пресмыкающихся, как и у земноводных, мезодерма разрастается между эктодермой и энтодермой, образуя характерные эмбриональные зачатки – сомиты, нефротомы и спланхнотомы.
Закладка осевых органов и других эмбриональных зачатков осуществляется в основном так же, как у других позвоночных. Образование кишечной трубки у пресмыкающихся во многом сходно с таковым у селяхий и костистых рыб в связи с наличием больших запасов желтка. Сворачивание слоя клеток энтодермы в кишечную трубку, образование желточного стебелька и разделение энтодермы на кишечную и желточную энтодерму происходит также при помощи туловищной складки. Туловищная складка у зародышей пресмыкающихся является принципиально новым образованием, возникающим в процессе эволюции, в связи с развитием амниотической оболочки.
Провизорные органы у зародышей пресмыкающихся возникают так же, как у зародышей хрящевых и костистых рыб. В связи с очень большим количеством желтка у них возникает желточный мешок. Образование желточного мешка у пресмыкающихся в отличие от рыб осуществляется значительно быстрее, и процесс обрастания желтка идет одновременно со всех сторон, тогда как у костистых рыб задний край бластодиска почти не принимает в этом участия. Если стенка желточного мешка у костистых рыб образована тремя зародышевыми листками, то у пресмыкающихся в связи с развитием амниотической оболочки эктодерма зародыша не принимает непосредственного участия в образовании стенки желточного мешка и образована лишь внезародышевой мезодермой висцерального листка спланхнотома и желточной энтодермой, т. е. двумя зародышевыми листками. У пресмыкающихся эктодерма и париетальный листок мезодермы образуют серозную оболочку, которая, располагаясь вокруг зародыша и его желточного мешка, предохраняет их от повреждений.
В процессе эмбриогенеза пресмыкающихся возникают три зародышевые оболочки: амнион, или водная оболочка, серозная, или наружная, оболочка и аллантоис, или мочевой мешок. Водная оболочка, возникает в процессе эволюции впервые у пресмыкающихся. Так как яйца пресмыкающихся не имеют белка, то развивающийся зародыш прилегает к плотным и упругим подскорлуповым оболочкам. Он лишен возможности, как это наблюдалось у хрящевых и костистых рыб, приподниматься над желтком в процессе обособления тела от околозародышевых частей бластодиска и погружается в разреженный желток. При этом зародыш пресмыкающихся прогибает слой клеток внезародышевой эктодермы, которая образует вокруг его тела складки, называемые амниотическими складками. Вначале в желток погружается передняя часть зародыша, а амниотические складки сходятся над ним и смыкаются.
Процесс смыкания амниотических складок распространяется постепенно на весь зародыш по мере его погружения в желток, в результате чего образуется амниотическая полость. Лишь на заднем конце зародыша амниотические складки не смыкаются и остается узкий серозно-амниотический канал, связывающий амниотическую полость с серозной полостью, т. е. пространством, ограниченным серозной оболочкой. Внешний листок эктодермы амниотической складки образует серозную оболочку, а внутренний – амниотическую оболочку.
Серозная и амниотическая оболочки пресмыкающихся состоят вначале лишь из эктодермы, а внезародышевая мезодерма, представленная париетальным листком спланхнотома, врастает позднее между наружным и внутренним листками внезародышевой эктодермы амниотической складки. Клетки эктодермы амниотической оболочки продуцируют амниотическую жидкость, которая омывает зародыш.
У пресмыкающихся при обрастании серозной оболочкой желточного мешка образуется промежуток между стенкой последнего и серозной оболочкой, который называют экзоцеломом или внезародышевым целомом. В экзоцелом врастает третья зародышевая оболочка – аллантоис, или мочевой мешок. У пресмыкающихся аллантоис образуется путем выпячивания нижней, или брюшной, стенки задней кишки зародыша. Стенка этого колбасовидного выпячивания состоит из двух зародышевых листков: кишечной энтодермы, которую снаружи покрывает висцеральный листок мезодермы спланхнотома. В мезодерме аллантоиса располагается густая сеть кровеносных сосудов, идущих от тела зародыша. Быстро разрастаясь в экзоцеломе, аллантоис подрастает к серозной оболочке, и его мезодерма срастается в местах контакта с мезодермой серозной оболочки. Впоследствии аллантоис заполняет все промежутки между серозной оболочкой, амнионом и желточным мешком. Аллантоис выполняет дыхательную и трофическую функции, переносит соли кальция. Основная функция аллантоиса – выделительная, поскольку в нем скапливаются конечные продукты обмена веществ в виде мочевой кислоты. В конце эмбрионального развития зародышевые оболочки редуцируются.
3. Эмбриогенез птиц
Развитие птиц в ряде случаев сходно с развитием пресмыкающихся. Дробление яйцеклетки птиц неполное, дискоидальное. При откладывании яйца зародыш находится на стадии бластулы или ранней гаструлы. В результате дробления образуется дискобластула. Первая фаза гаструляции у птиц осуществляется путем деляминации, при которой возникают эктодерма и энтодерма (рисунок 2.2).
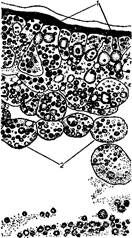
1 – клетки эктодермы; 2 – клетки энтодермы
Рисунок 2.2 – Первая фаза гаетруляции у птиц
Вторая фаза гаструляции у птиц отлична от таковой у пресмыкающихся. Бластодиск зародыша птиц очень разрастается, и в центре его возникает зародышевый щиток, клеточный материал которого идет на построение тела. Клеточный материал зародышевого щитка птиц в связи с интенсивным размножением клеток эктодермы начинает двигаться двумя потоками, справа и слева от средней сагиттальной линии к его заднему краю. Это движение клеточного материала ведет к удлинению первоначально круглого зародышевого щитка. На заднем конце зародышевого щитка клеточные потоки встречаются, поворачивают и начинают двигаться вперед по средней сагиттальной линии, образуя в этом месте утолщенный слой клеток, называемый первичной полоской. Передняя часть первичной полоски образует расширение, называемое первичным или гензеновским узелком. Встречные потоки клеточного материала в области первичного узелка образуют углубление, называемое первичной ямкой. Посередине первичной полоски возникает продольное углубление – первичная бороздкая или первичный желобок. Он является продолжением назад первичной ямки. Первичная ямка представляет собой верхнюю, или спинную, губу, первичная бороздка – боковые губы, а конец первичной бороздки является нижней, или брюшной, губой бластопора (рисунок 2.3).
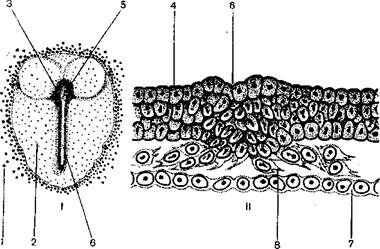
I – зародышевый диск (вид сверху); II – образование мезодермы; 1 – желток;
2 – зародышевый диск; 3 – гензеновский узелок; 4 – эктодерма; 5 – первичная ямка;
6 – первичная полоска; 7 – энтодерма; 8 – клетки мезодермы
Рисунок 2.3 – Вторая фаза гаструляции у птиц (ориг.)
Образование мезодермы во второй фазе гаструляции осуществляется путем инвагинации клеточного материала эктодермы через первичную ямку и первичную бороздку в пространство между наружным и внутренним зародышевыми листками. Клеточный материал хордальной пластинки, располагающийся впереди от первичной ямки, начинает подворачиваться и инвагинировать через первичную ямку вперед и вниз под эктодерму, формируя головной, или хордальный, отросток, который растет между эктодермой и энтодермой. Через переднюю часть первичной бороздки начинает инвагинировать вглубь, вперед и в стороны клеточный материал сегментированной мезодермы эктодермального происхождения. Сегментированная мезодерма, располагаясь по бокам хорды, дает начало сомитам. Клеточный материал, который инвагинирует в задней части первичной бороздки и является производным эктодермы, дает начало несегментированной мезодерме.
Кроме образования мезодермы путем инвагинации у зародышей птиц в области первичной бороздки, наблюдается вселение, или иммиграция, клеточного материала мезодермы из эктодермы. По мере инвагинации клеточного материала хорды и мезодермы первичный узелок и первичная ямка постепенно перемещаются назад к брюшной губе бластопора, ввиду чего первичная полоска укорачивается и первичный узелок оказывается у брюшной губы бластопора, на заднем конце зародыша.
По окончании второй фазы гаструляции зародыш птиц становится трехслойным. Часть эктодермы, которая располагается впереди от первичного узелка, над хордой, дает начало нервной пластинке, а вся остальная эктодерма – эпителию кожи. Закладка осевых органов и других эмбриональных зачатков у зародышей птиц идет так же, как у пресмыкающихся.
В образовании провизорных органов птиц много общего с этими процессами у пресмыкающихся. Желточный мешок у зародышей птиц возникает так же, как у зародышей пресмыкающихся. В процессе эмбриогенеза у зародышей птиц появляются три зародышевые оболочки: амниотическая, серозная и аллантоис. Образование амниотической и серозной оболочек у птиц отличается от таковых у пресмыкающихся, так как между зародышем и подскорлуповой оболочкой яйца птиц располагается толстый слой белка. Поэтому развивающийся зародыш не погружается в желток, а приподнимается над ним. Вследствие неравномерного роста краев бластодиска по обеим сторонам зародыша возникает углубление, называемое туловищной складкой, в состав которой входят внезародышевая эктодерма и париетальный листок спланхнотома внезародышевой мезодермы (рисунок 2.4).
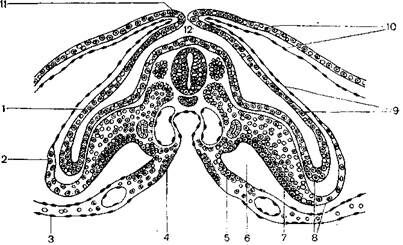
1– эктодерма; 2 – мезодерма; 3 – желточная энтодерма; 4 – кишечная энтодерма; 5 – висцеральный листок спланхнотома; 6 – целом; 7 – париетальный листок спланхнотома; 8 – туловищная складка; 9 – будущая амниотическая оболочка; 10 – будущая серозная оболочка; 11 – амниотическая складка;
12 – будущая амниотическая полость.
Рисунок 2.4 – Образование зародышевых оболочек у птиц
Углубляясь и разрастаясь, туловищная складка приподнимает зародыш над желтком и способствует сворачиванию в трубку кишечной энтодермы. Туловищная складка продолжается в амниотические складки, которые поднимаются над зародышем и срастаются, образуя амниотическую полость. Срастание амниотических складок осуществляется послойно: внутренний листок эктодермы срастается с таковым противоположной стороны, наружный – с наружным, то же самое происходит и с двумя листками мезодермы. В результате срастания амниотических складок возникают одновременно две зародышевые оболочки: внутренняя – амниотическая и наружная – серозная.
В амниотической оболочке эктодермальный слой клеток обращен внутрь, к зародышу, а мезодермальный – наружу, а в серозной оболочке, наоборот, мезодермальный слой обращен внутрь, к мезодерме амниотической оболочки, а эктодермальный слой – наружу – к белку яйца. Серозная оболочка, разрастаясь, образует наружную зародышевую оболочку, которая обрастает снаружи не только зародыш и желток, но и ту часть белка, которая в процессе развития остается лишь у острого конца яйца под дном желточного мешка. Клетки серозной оболочки эктодермального происхождения, которые контактируют с белком, принимают участие в его резорбции при помощи многочисленных микроворсинок. Дальнейший путь питательных веществ идет по сосудам мезодермы аллантоиса к телу зародыша. Таким образом, одной из функций серозной оболочки является трофическая. Часть белка яйца по серозно-амниотическому каналу поступает в амниотическую полость и заглатывается плодом. Кроме трофической функции, серозной оболочке присуща функция дыхания.
В процессе развития зародыша и потребления им белка серозная оболочка разрастается и контактирует с подскорлуповой оболочкой яйца на всем протяжении, в том числе и в области воздушной камеры. Сосуды мезодермы аллантоиса, которые врастают в мезодерму серозной оболочки, вместе с эпителием серозной оболочки образуют барьер «кровь – воздух» и осуществляют функцию газообмена. Кроме того, серозная оболочка принимает участие в минеральном обмене. Клетки поверхностного слоя серозной оболочки имеют многочисленные микроворсинки, которые тесно контактируют с сосочками скорлупы яйца. Эти клетки богаты митохондриями и секретируют соляную кислоту. Соляная кислота вводится в канальцы скорлупы яйца и растворяет соли кальция, которые через клетки серозной оболочки попадают в кровяное русло мезодермы серозной оболочки и аллантоиса и переносятся в тело зародыша для построения его скелета. Аллантоис у зародышей птиц развивается так же, как у зародышей пресмыкающихся, и выполняет те же функции.
4. Эмбриогенез млекопитающих
В связи с некоторыми особенностями эмбрионального развития млекопитающих животных, относящихся к отряду приматов, а также человека их эмбриогенез рассматривается отдельно от других систематических групп млекопитающих.
Эмбриональное развитие яйцекладущих млекопитающих имеет много общего с развитием пресмыкающихся и птиц, так как яйцеклетки утконоса и ехидны очень богаты желтком и имеют сходное строение. Развитие зародышей у этих видов млекопитающих идет во время насиживания или при вынашивании яйца в сумке, т. е. во внешней среде. Дробление яйцеклетки неполное, дискоидальное, с образованием дискобластулы. Гаструляция, закладка осевых органов, развитие зародышевых оболочек осуществляется почти так же, как у пресмыкающихся.
У сумчатых млекопитающих яйцеклетки мезолецитальные, по расположению желтка их можно отнести к телолецитальным яйцеклеткам, поскольку желток локализован в области вегетативного полюса. Дробление яйцеклетки полное, неравномерное, лишь на вегетативном полюсе остается нераздробленным незначительное количество желтка (рисунок 3.1).
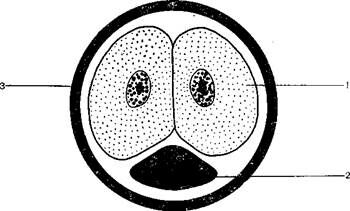
1 – бластомер; 2 – желток; 3 – вторичная оболочка яйцеклетки
Рисунок 3.1 – 5. Дробление яйцеклетки сумчатой куницы:
На начальных стадиях дробления бластомеры располагаются в анимально-экваториальной части над желточным шаром и полностью отделены от желтка. При последующих дроблениях бластомеры занимают экваториальное положение, а желточный шар располагается в центре. С увеличением количества бластомеров они постепенно достигают полюсов, полностью обрастая желток, который оказывается в центре бластодермического пузырька, или бластоцисты, где находится уже в жидком виде.
В связи с незначительным количеством желтка в яйцеклетке процесс его обрастания у сумчатых млекопитающих проходит быстрее, чем у пресмыкающихся, птиц и яйцекладущих млекопитающих. В однослойном бластодермическом пузырьке клетки, расположенные на анимальном полюсе, несколько мельче клеток, находящихся на вегетативном полюсе. Клеточный материал анимального полюса бластоцисты является исходным для образования зародыша. Стенка бластодермического пузырька у анимального полюса путем деляминации и иммиграции отщепляет в полость бластоцисты слой клеток будущей энтодермы, а сама превращается в наружный зародышевый листок – эктодерму. На этом завершается первая фаза гаструляции.
Далее возникает первичный узелок и первичная полоска, и происходит вторая фаза гаструляции с образованием мезодермы путем инвагинации эктодермы с последующей иммиграцией клеток в пространство между эктодермой и энтодермой. Эта фаза гаструляции, а также закладка осевых органов проходят примерно так же, как у пресмыкающихся, птиц и яйцекладущих млекопитающих.
Развитие сумчатых млекопитающих имеет следующие особенности:
1) клетки внезародышевой бластодермы, т. е. наружные стенки бластодермического пузырька, образуют трофобласт – слой клеток, который, прилегая к слизистой оболочке матки, приобретает способность к всасыванию питательных веществ – так называемого маточного молочка, выделяемого железами слизистой оболочки матки.
2) возникновение плаценты примитивного типа, которая получила название полуплаценты. Этот орган является принципиально новым в процессе эволюции и формируется он не у всех сумчатых. Полуплацента представляет собой серозную оболочку, переходящую в трофобласт, на поверхности которой клетки эктодермы образуют простые неразветвленные ворсинки. Ворсинки серозной оболочки и трофобласта проникают в отверстия маточных желез, но не достигают соединительнотканной основы слизистой оболочки матки. С образованием первичных ворсинок серозная оболочка и трофобласт сумчатых превращаются в хорион, или ворсинчатую оболочку, осуществляющую связь зародыша с организмом матери.
У плацентарных млекопитающих наблюдается вторичная утрата желтка яйцеклеткой в связи с внутриутробным развитием зародышей. Яйцеклетки у этих животных олиголецитальные, дробление полное, неравномерное. Дробление яйцеклетки происходит во время ее продвижения по яйцеводу. В это время дробящаяся яйцеклетка у некоторых животных, например у грызунов, зайцеобразных, хищных, непарнопалых, покрывается довольно толстым слоем белка, который секретируют клетки яйцевода.
Первая борозда дробления проходит меридионально. Второе деление происходит спустя два-три дня и также проходит меридионально, третье дробление идет в экваториальной плоскости. Начиная со второго дробления бластомеры начинают делиться несинхронно, и после второго дробления зародыш представлен лишь тремя бластомерами. У некоторых млекопитающих уже после первых дроблений яйцеклетки наблюдается различие в строении бластомеров: одни бластомеры более темные, а другие имеют светлую, прозрачную цитоплазму. Темные бластомеры называют зародышевыми или базофильными, их цитоплазма имеет большое количество РНК. Эти бластомеры располагаются у анимального полюса морулы. Бластомеры со светлой цитоплазмой называют внезародышевыми, они располагаются у вегетативного полюса. Эта ранняя дифференциация бластомеров показывает, что темные бластомеры являются исходным материалом для образования тела зародыша, а светлые – для образования трофобласта (рисунок 3.2).
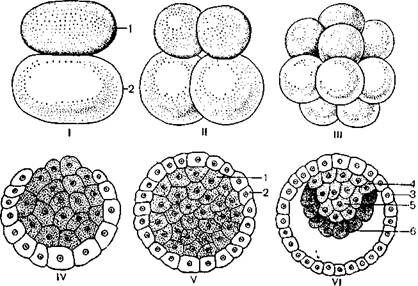
I – стадия 2 бластомеров; II – стадия 4 бластомеров; III – морула; IV–V – образование трофобласта; VI – бластоциста и первая фаза гаструляции:
1 – темные бластомеры; 2 – светлые бластомеры; 3 – трофобласт;
4 – эмбриобласт; 5 – эктодерма; 6 – энтодерма.
Рисунок 3.2 – Начальные стадии эмбриогенеза млекопитающих животных
После четвертого дробления образуется морула и наблюдается перегруппировка бластомеров: один из них, более темный, смещается от анимального полюса к центру, а три темных и четыре светлых бластомера располагаются вокруг. Потом в центр перемещается еще один темный бластомер, а остальные располагаются по периферии. Начиная с этой стадии развития дробление бластомеров делается несинхронным: светлые, внезародышевые бластомеры делятся быстрее, чем темные, зародышевые бластомеры. В результате неравномерного дробления внезародышевые бластомеры наползают на зародышевые и постепенно обрастают их. С завершением обрастания темные бластомеры, или клетки эмбриобласта, оказываются внутри, а снаружи располагаются светлые бластомеры, или клетки трофобласта. В результате жизнедеятельности клеток трофобласта между ними и клетками эмбриобласта возникает полость и образуется бластоциста (бластодермический, или зародышевый, пузырек). Эмбриобласт формирует зародышевый узелок, а трофобласт – временную оболочку, которая обеспечивает питание зародыша до возникновения иных зародышевых оболочек.
Все описанные процессы осуществляются в зародыше плацентарных млекопитающих в то время, когда он представляет собой изолированную систему и не находится в обменных взаимоотношениях с материнским организмом, так как от него он отделен остатками вторичной оболочки – ее светлой зоной. С распадом остатков вторичной оболочки яйцеклетки трофобласт вступает в тесный контакт со слизистой оболочкой матки, улучшается питание зародыша и клетки бластоцисты начинают усиленно размножаться. На наружной поверхности клеток трофобласта, там, где располагается эмбриобласт, образуются выросты – ворсинки, которые погружаются в стенку матки. Этот процесс называют имплантацией. Имплантация зародыша в стенку матки у млекопитающих различных систематических групп наступает в различные сроки после оплодотворения – от первой до десятой недели. Впоследствии трофобласт соединяется с серозной оболочкой и входит в состав детского места, или плаценты.
Гаструляция у плацентарных млекопитающих протекает так же, как у пресмыкающихся, птиц и низших млекопитающих. От поверхности зародышевого узелка, которая обращена в полость зародышевого пузырька, отщепляется путем деляминации слой плоских клеток, представляющий собой энтодерму, а вся остальная клеточная масса зародышевого узелка именуется эктодермой. Энтодерма разрастается по всей внутренней поверхности трофобласта, образуя энтодермальную стенку первичного желточного мешка. Ту часть энтодермы, которая контактирует с клетками трофобласта, называют внезародышевой энтодермой, а ту, что граничит с эктодермой – зародышевой энтодермой.
После образования двуслойного зародыша клетки эктодермы растягиваются и образуют зародышевый щиток, который непосредственно продолжается в трофобласт. Трофобласт над зародышевым щитком истончается и разрывается, вследствие чего до образования амниона зародыш млекопитающих ничем не покрыт. В течение второй фазы гаструляции образуется мезодерма так же, как у птиц и низших млекопитающих, и зародыш становится трехслойным. Закладка осевых органов принципиально ничем не отличается от таковой у птиц и низших млекопитающих.
Зародышевые оболочки у плацентарных млекопитающих возникают в основном так же, как у пресмыкающихся, птиц и низших млекопитающих, однако в деталях строения, функции и возникновения некоторых зародышевых оболочек наблюдаются существенные различия (рисунок 3.3).
Во-первых, при возникновении хориона у млекопитающих, как и у птиц, вначале образуются примитивные первичные ворсинки трофобласта, которые затем сменяются сложными вторичными ворсинками хориона. Вторичные ворсинки возникают вследствие подрастания к хориону аллантоиса, срастания их мезодермы и врастания кровеносных сосудов аллантоиса в ворсинки хориона. С врастанием кровеносных сосудов ворсинки хориона становятся крупными, ветвистыми, увеличивая площадь соприкосновения зародышевых оболочек с организмом матери.
Во-вторых, зародышевые оболочки плацентарных млекопитающих отличаются от оболочек птиц тем, что аллантоис млекопитающих не является мочевым мешком, поскольку мочевина из тела зародыша выделяется через плаценту в кровь матери.
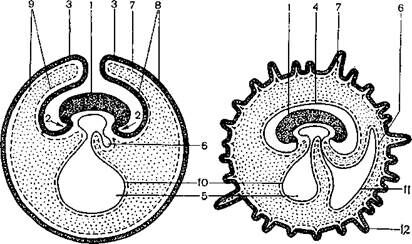
I – начальные стадии образования амниона, аллантоиса и серозной оболочки; II – образование амниона, аллантоиса и хориона с первичными ворсинками;
1 – тело зародыша; 2 – туловищные складки; 3 – амниотические складки; 4 – амниотическая оболочка; 5 – желточный мешок; 6 – аллантоис; 7 – сросшиеся серозная оболочка и трофобласт – хорион; 8 – эктодерма; 9 – мезодерма; 10 – желточная энтодерма; 11 – энтодерма аллантоиса; 12 – первичные ворсинки хориона.
Рисунок 3.3 – Развитие зародышевых оболочек у плацентарных млекопитающих
Часть аллантоиса млекопитающих принимает затем участие в образовании стенки мочевого пузыря. Связь между телом зародыша и плацентой осуществляется при помощи пуповины, или пупочного канатика, который состоит из протока желточного мешка, аллантоиса и его сосудов – пупочной вены, двух пупочных артерий. Снаружи пупочный канатик покрыт амниотической оболочкой (рисунок 3.4).
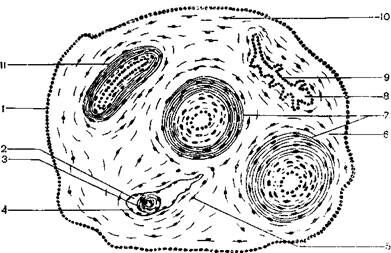
1 – эктодерма амниотической оболочки; 2 – желточный мешок; 3 – желточный стебелек; 4 – желточная энтодерма; 5 – мезодерма желточного мешка; 6 – мезодерма аллантоиса; 7 – пупочные артерии; 8 – аллантоис; 9 – энтодерма аллантоиса; 10 – мезодерма амниотической оболочки; 11 – пупочная вена.
Рисунок 3.4 – Пупочный канатик млекопитающего
В-третьих, у некоторых плацентарных млекопитающих, например рукокрылых, насекомоядных, приматов, амнион возникает не путем образования амниотических складок, а в результате появления полости между клетками зародышевого узелка, т. е. его образование смещается на более ранние стадии эмбрионального развития (стадия бластоцисты), еще до образования зародышевых листков. У перечисленных млекопитающих при таком способе образования амниона нет серозной оболочки, и в образовании плаценты принимает участие трофобласт, его производное – хорион, а также аллантоис.
5. Эмбриогенез человека и приматов
Эмбриогенез человека и приматов несколько отличается от развития других плацентарных млекопитающих. Первые стадии дробления яйцеклеток у человека и остальных приматов происходят так же, как и у других плацентарных млекопитающих. У человека в течение первых суток после оплодотворения яйцеклетка делится на два почти одинаковых по величине бластомера. Дробление яйцеклетки полное, неравномерное. Еще через сутки в результате второго дробления возникают 3–4 бластомера. Спустя четверо суток после оплодотворения зародыш человека образует 8–12-клеточную морулу (рисунок 3.5).
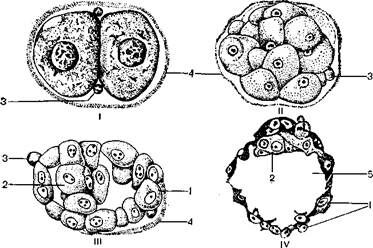
I – стадия 2 бластомеров; II – морула; III – ранняя бластоциста; IV – 107-клеточная
бластоциста;
1 – трофобласт; 2 – эмбриобласт; 3 – направительные (полярные) тельца; 4 – остатки блестящей зоны вторичной оболочки яйцеклетки; 5 – полость бластоцисты.
Рисунок 3.5 – Начальные стадии эмбриогенеза человека:
Во время дробления яйцеклетки человека наблюдается образование темных и светлых бластомеров, т. е. так же, как и у других плацентарных млекопитающих. Начиная со стадии 12-клеточной морулы светлые бластомеры будущего трофобласта начинают обрастать темные, базофильные бластомеры будущего эмбриобласта. В результате обрастания и накопления клетками трофобласта белковой жидкости между клетками зародыша образуется неправильной формы полость и возникает ранняя бластоциста, у которой клетки эмбриобласта располагаются в центре, а клетки трофобласта – по периферии. У человека на четвертые сутки после оплодотворения зародыш попадает в полость матки; это приходится на 18–19-й день менструального цикла. Весь период развития – от оплодотворения и до попадания зародыша в полость матки вокруг него сохраняются остатки вторичной оболочки яйцеклетки. В это время питание зародыша осуществляется за счет собственных запасов желтка.
Наличие остатков вторичной оболочки яйцеклетки вокруг развивающегося зародыша препятствует обменным процессам с организмом матери, преждевременному прикреплению зародыша к слизистой оболочке маточной трубы и наступлению внематочной беременности. Когда зародыш человека попадает в полость матки, остатки вторичной оболочки яйцеклетки разрушаются, и он получает возможность питаться маточным молочком – секретом желез слизистой оболочки матки. С переходом на питание маточным молочком развитие зародыша ускоряется. К середине шестых суток он уже состоит из 107 клеток и превращается в зрелую бластоцисту. Стенка зрелой бластоцисты состоит из плоских, удлиненных клеток, называемых первичным цитотрофобластом. К цитотрофобласту изнутри прилегает скопление клеток, образующих эмбриобласт, или зародышевый узелок, т. е. клеточный материал, из которого образуется тело зародыша. Место расположения клеток зародышевого узелка в бластоцисте назывется эмбриональным полюсом бластоцисты.
У человека и приматов амнион возникает на ранних стадиях эмбрионального развития, еще до образования зародышевых листков и закладки осевых органов. В результате уплощения клеток эмбриобласта образуется зародышевый диск, гомологичный бластодиску пресмыкающихся и птиц. В начале седьмых суток после оплодотворения у зародыша человека наблюдается перемещение клеток зародышевого щитка, в результате чего между клетками змбриобласта возникает полость. Такой тип образования полости внутри клеточной массы называют кавитацией. Эта полость является амниотической полостью.
К исходу седьмых суток после оплодотворения наступает первая фаза гаструляции. Клетки зародышевого узелка путем деляминации отщепляют в сторону полости бластоцисты тонкий слой клеток энтодермы, а остальная масса клеток зародышевого узелка представляет собой эктодерму. Таким образом, эктодерма и энтодерма зародыша возникают еще до имплантации. К восьмому дню внутриутробного развития человека бластоциста прикрепляется к слизистой оболочке матки и начинается процесс имплантации (20–21-й день менструального цикла).
После прикрепления к задней стенке матки эмбриональным полюсом бластоциста начинает погружаться в слизистую оболочку матки и наступает имплантация. К исходу вторых суток от начала имплантации зародыш человека полностью погружается в слизистую оболочку матки (рисунок 3.6).
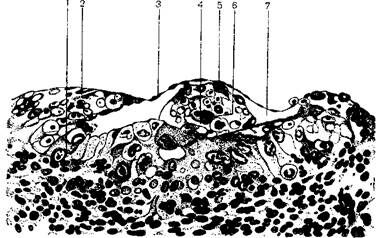
1 – трофобласт; 2 эпителий матки; 3 – стенка бластоцисты; 4 – полость амниона; 5 – энтодерма и 6 – эктодерма зародыша; 7 – полость бластоцисты
Рисунок 3.6 – Зародыш человека 7,5 дней в периоде имплантации и образования амниотической полости
Процесс имплантации осуществляется благодаря деятельности клеток цитотрофобласта. С началом имплантации клетки цитотрофобласта эмбрионального полюса начинают усиленно размножаться и часть клеток, прилегающих к матке, дифференцируется в клетки плазмодиотрофобласта. Эти клетки отличаются от клеток цитотрофобласта тем, что 1) имеют очень большие ядра с несколькими крупными ядрышками; 2) размеры этих клеток возрастают во много раз, образуя многочисленные отростки разнообразной формы; 3) клетки имеют резко базофильную цитоплазму, обусловленную большим содержанием РНК и хорошо развитой гранулярной эндоплазматической сетью. Клетки плазмодиотрофобласта образуют синцитий – сетевидное скопление отдельных клеток, в связи с чем они называются синцитиальным трофобластом.
Клетки имеют резко выраженные ферментативные свойства, активно растворяют эпителий матки и глубоко внедряются в отечную соединительнотканную основу слизистой оболочки матки. Продукты распада слизистой оболочки матки – эмбриотроф – фагоцитируются клетками трофобласта и используются зародышем для питания. В результате деятельности протеолитических ферментов плазмодиотрофобласта в слизистой оболочке матки возникают небольшие полости, при слиянии которых на девятый день после оплодотворения образуются лакуны трофобласта. Лакуны трофобласта вначале не связаны между собой, но затем клетки плазмодиотрофобласта начинают разрушать стенки между лакунами, а также стенки венозных синусоидов слизистой оболочки матки, и лакуны трофобласта заполняются материнской кровью. С десятого дня развития зародыша человека протеолитическая активность клеток плазмодиотрофобласта начинает затухать, клетки дегенерируют, синцитий распадается на фрагменты, которые в дальнейшем постепенно исчезают.
Клетки цитотрофобласта, бурно размножаясь, внедряются в слизистую оболочку матки и замещают гигантские клетки плазмодиотрофобласта. Скопления клеток цитотрофобласта дают начало первичным ворсинкам трофобласта, в которых не наблюдается еще кровеносных сосудов. В первичные ворсинки трофобласта на 12-й день развития зародыша врастают клетки внеэмбриональной, или внезародышевой, мезенхимы, которые образуются у зародыша человека на 9-й день развития в результате отщепления клеток от трофобласта. Размножающиеся клетки внезародышевой мезенхимы покрывают изнутри поверхность бластоцисты и осуществляют связь между зародышем и трофобластом. С врастанием внезародышевой мезенхимы первичные ворсинки трофобласта начинают увеличиваться в размерах, ветвиться, однако в них еще нет кровеносных сосудов.
К концу второй недели развития первичные ворсинки трофобласта превращаются во вторичные, которые отличаются от первичных тем, что основу их образуют клетки внезародышевой мезенхимы.
Далее кнаружи располагаются клетки цитотрофобласта, а за ними – клетки видоизмененного плазмодиотрофобласта (рисунок 3.7).
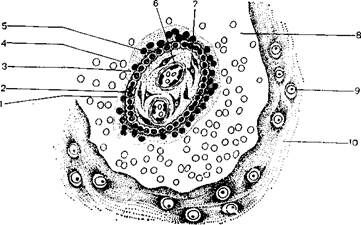
1 – поперечный срез ворсинки хориона; 2 – цитотрофобласт; 3 – плазмодиотрофобласт (синцитиальный трофобласт); 4 – эритроциты крови матери; 5 – ядра клеток плазмодиотрофобласта; 6 – кровеносные сосуды ворсинки; 7 – мезенхима ворсинки;
8 – гемохориальное пространство, заполненное кровью матери;
9 – клетки периферического цитотрофобласта и отпадающие клетки слизистой оболочки матки; 10 – фибриноид
Рисунок 3.7 – Ворсинка хориона человека
С образованием вторичных ворсинок трофобласт с подстилающей его внезародышевой мезенхимой превращается в ворсинчатую оболочку, или хорион.
Параллельно с имплантацией идет развитие самого зародыша. После завершения первой фазы гаструляции и образования двухслойного зародыша эктодерма и энтодерма включает в себя клеточный материал самого зародыша и внезародышевый клеточный материал, из которого в дальнейшем возникнут стенки амниона и желточного мешка. Дно амниона, обращенное к клеткам энтодермы, контактирует с эктодермой зародыша, а остальная эктодерма является стенкой и крышей амниона. Энтодерма, граничащая с эктодермой зародыша, является зародышевой энтодермой. Зародышевая эктодерма и энтодерма именуются зародышевым щитком.
К 12-му дню развития зародыша человека клетки энтодермы разрастаются и под зародышевым щитком образуют первичный желточный мешок, который, как и амнион, имеет вид пузырька. В этот период развития зародыш состоит из двух неодинаковых по величине пузырьков, которые, соприкасаясь между собой, образуют двухслойный зародышевый щиток, являющийся материалом для развития тела зародыша. Меньший из этих пузырьков, обращенный к стенке матки, называется амниотическим, а больший – первичным желточным пузырьком. Снаружи оба пузырька окружены слоем клеток внезародышевой мезодермы, которые в одном лишь месте – в будущем заднем конце зародыша – переходят на внутреннюю поверхность трофобласта, образуя поддерживающий стебелек, или амниотическую ножку. В амниотическую ножку в дальнейшем будет врастать аллантоис, соединяясь с хорионом.
На 13–14-й день после оплодотворения энтодерма зародыша сворачивается в кишечную трубку, а внезародышевая эктодерма образует постоянный, или дефинитивный, желточный мешок. Энтодерма первичной кишки зародыша остается связанной с внезародышевой энтодермой желточного мешка, или желточной энтодермой, лишь в одном месте при помощи широкого желточного протока. Внезародышевая мезодерма вначале заполняет всю полость зародышевого пузырька, в дальнейшем между клетками мезенхемы возникают небольшие полости, наполненные жидкостью, при слиянии которых образуется полость, расположенная вокруг зародыша – экзоцелом, или внезародышевый целом (рисунок 3.8).
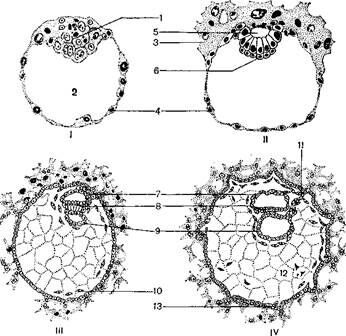
I – 6-дневная бластоциста; II – 8-дневный зародыш, первая фаза гаструляции;
III – 12-дневный зародыш; IV – 14-дневный зародыш;
1 – эмбриобласт; 2 – полость бластоцисты; 3 – трофобласт; 4 – стенка бластоцисты; 5 – амниотическая полость; 6 – энтодерма зародыша; 7 – эктодерма амниона; 8 – эктодерма зародыша; 9 – желточная энтодерма;
10 – внезародышевая мезодерма; 11 – амниотическая ножка; 12 – внезародышевый целом; 13 – первичные ворсинки хориона
Рисунок 3.8 – Эмбриогенез человека
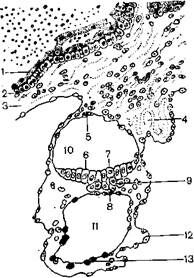
1 – плазмодиотрофобласт; 2 – цитотрофобласт; 3 – соединительная ткань хориона;
4 – амниотическая ножка; 5 – эктодерма амниона; 6 – зародышевый щиток;
7 – эктодерма зародыша; 8 –энтодерма зародыша; 9 – мезодерма зародыша;
10 – амниотическая полость; 11 – полость желточного мешка;
12 – внезародышевая мезодерма; 13 – желточная энтодерма
Рисунок 3.9 – Вторая фаза гаструляции 15-дневного зародыша человека
К этому же периоду у зародыша наблюдается формирование еще одной зародышевой оболочки – аллантоиса. На 15-й день после оплодотворения задний отдел кишечника образует сначала пальцевидный, а затем колбасовидный вырост. Стенка этого выроста представлена энтодермой первичной кишки, поверх которой располагается мезодерма висцерального листка спланхнотома. Аллантоис врастает в мезодерму амниотической ножки в направлении трофобласта, а сама ножка превращается в аллантоидную ножку. Значение аллантоиса в питании и дыхании зародыша велико, так как по мезодерме аллантоиса растут к хориону кровеносные сосуды, которые разветвляются во вторичных ворсинках.
По окончании гаструляции зародыш человека и других приматов переходит в следующий, период развития, длящийся с 21-го по 35-й день, в течение которого осуществляется нейруляция и закладка осевых органов, происходящая примерно так же, как и у других плацентарных млекопитающих.
В питании, дыхании и обмене веществ зародыша важнейшую роль играет плацента, или детское место. Плацента, как орган связи зародыша с материнским организмом, свойственна не только плацентарным и некоторым сумчатым млекопитающим, но и ряду беспозвоночных, например членистоногим, из рыб – живородящим акулам. Однако лишь у плацентарных млекопитающих животных и человека строение плаценты наиболее сложно и совершенно. В образовании плаценты у человека и прочих приматов принимают участие аллантоис, хорион зародыша и слизистая оболочка матки. Хорион зародыша вначале на всем протяжении равномерно покрыт ворсинками. Основа каждой ворсинки состоит из внезародышевой мезенхимы, из которой в дальнейшем развивается рыхлая, неоформленная соединительная ткань, переходящая в соединительнотканную основу хориона. Ворсинки хориона покрыты клетками цитотрофобласта, на поверхности которых располагается плазмодиотрофобласт.
На 18-й день после оплодотворения в соединительнотканной основе ворсинок хориона зародыша человека появляются зачатки кровяных островков и образуются первые кровеносные сосуды. К этому периоду аллантоис подрастает к хориону и мезодерма последнего срастается с таковой аллантоиса. Кровеносные сосуды зародыша, расположенные в мезодерме аллантоиса, врастают в соединительнотканную основу хориона и в ворсинки последнего. На 20–21-й день после оплодотворения сосуды ворсинок хориона соединяются с кровеносными сосудами зародыша.
Несмотря на установление прямой связи между кровеносными сосудами зародыша и хориона, эта система начинает функционировать лишь к 23–24-му дню после оплодотворения, т. е. тогда, когда у зародыша начинает сокращаться сердце и кровь через пупочные артерии начинает поступать в кровеносные сосуды ворсинок хориона. С этого момента устанавливается связь между кровеносной системой ворсинок хориона и кровеносной системой организма матери. Однако между двумя кровеносными системами нет прямой связи, так как между ними располагаются клеточные элементы стенки ворсинки хориона – плазмодиотрофобласт, цитотрофобласт, а также стенки кровеносных сосудов хориона плода.
Связь тела зародыша с плацентой осуществляется при помощи пупочного канатика, основой которого сначала является поддерживающий стебелек, преобразующийся впоследствии в амниотическую ножку, а затем в аллантоидную ножку. По мере роста амниона его стенки постепенно покрывают аллантоидную ножку, в которой проходят аллантоис, желточный проток, две пупочные артерии и одна пупочная вена. Мезодерма указанных образований срастается и одевается снаружи мезодермой и эктодермой амниона.
6. Типы плацент
В связи с особенностями проникновения ворсинок хориона зародыша в слизистую оболочку матки плаценты млекопитающих подразделяют на 4 типа: эпителиохориальную, десмохориальную, эндотелиохориальную и гемохориальную плаценту (рисунок 3.10).
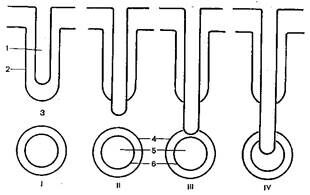
I – эпителиохориальная; II – десмохориальная; III – эндотелиохориальная; IV – гемо-хориальная;
1 – ворсинка хориона; 2 – эпителий маточной крипты; 3 – соединительная ткань слизистой оболочки матки; 4 – стенка кровеносной сосудистой системы слизистой оболочки матки; 5 – просвет кровеносной системы;
6 – эндотелий кровеносной системы
Рисунок 3.10 – Типы плацент
Плацента эпителиохориального типа, или полуплацента, свойственна некоторым сумчатым, например сумчатому барсуку, и плацентарным млекопитающим: свинье, лошади, верблюду, бегемоту, дельфинам и пр. Плацента этого типа характеризуется тем, что ворсинки хориона зародыша только соприкасаются с эпителием матки. Ворсинки хориона лишь входят в углубления слизистой оболочки матки – отверстия маточных желез и не разрушают их. Питательные вещества и кислород поступают в кровеносную систему зародыша из маточного молочка – секрета желез слизистой оболочки матки. При родах ворсинки хориона вытягиваются из отверстий маточных желез слизистой оболочки матки и разрушения последней не наблюдается.
Десмохориальная, или соединительнотканнохориальная, плацента свойственна парнопалым жвачным. Плацента этого типа характеризуется тем, что ворсинки хориона зародыша погружаются в эпителий матки, разрушают его и проникают в подлежащую соединительную ткань. Таким образом, ворсинки хориона зародыша располагаются уже ближе к кровяному руслу матки. При этом связь зародыша с организмом матери также не очень тесная, однако это компенсируется значительным увеличением поверхности хориона. Во время родов на поверхности слизистой оболочки матки после отделения плаценты остаются участки, лишенные эпителия. Эпителиальный слой слизистой оболочки матки быстро регенерирует за счет смежных участков неповрежденного эпителия.
Эндотелиохориальная, или вазохориальная, плацента свойственна хищным. В эндотелиохориальной плаценте ворсинки хориона зародыша, погружаясь в стенку матки, разрушают эпителий, подлежащую соединительную ткань и стенку кровеносных сосудов матки до эндотелиального слоя. Ворсинки хориона зародыша контактируют с эндотелиальным слоем клеток кровеносных сосудов матки, и лишь тончайший слой эндотелиальных клеток отделяет ворсинки хориона от потока крови в сосудах матки. Этот способ образования плаценты обеспечивает лучшее снабжение зародыша кислородом и питательными веществами, а также ведет к значительному уменьшению величины плаценты. При родах часть тканей слизистой оболочки матки отторгается и возникает небольшое кровотечение, а дефекты слизистой оболочки быстро ликвидируются.
Гемохориальная плацента свойственна человеку и другим приматам, а также некоторым насекомоядным, рукокрылым, грызунам. Этот тип плаценты характеризуется тем, что ворсинки хориона зародыша прорастают эпителий, подлежащую соединительную ткань, стенки кровеносных сосудов слизистой оболочки матки и ворсинки хориона омываются кровью матери.
Гемохориальная плацента может быть лабиринтовой и ворсинчатой. Лабиринтовая гемохориальная плацента присуща насекомоядным и грызунам. У этой плаценты трофобласт образует систему полостей, которые сливаются в сложный лабиринт, по которому течет материнская кровь. Ворсинчатая гемохориальная плацента имеется у человека и прочих приматов. Ворсинки хориона у плаценты этого типа сильно ветвятся, образуя тем самым колоссальную поверхность, и омываются кровью матери. При родах разрушается вся отпадающая оболочка слизистой оболочки матки и возникает сильное кровотечение. Слизистая оболочка матки ввиду значительного дефекта регенерирует на протяжении продолжительного времени.
Вопросы для закрепления.
1 Каких животных относят к амниотам?
2. Когда происходит дробление яйцеклетки у пресмыкающихся?
3. Опишите первую фазу гаструляции у пресмыкающихся.
4. Дайте характеристику второй фазы гаструляции у пресмыкающихся.
5. Что называется у пресмыкающихся первичной пластинкой, как она образуется?
6. Как происходит у пресмыкающихся образование полости первичной кишки?
7. Охарактеризуйте процесс закладки осевых органов у пресмыкающихся.
8. Как происходит образование желточного мешка у пресмыкающихся, какие функции он выполняет?
9. Перечислите зародышевые оболочки пресмыкающихся, как они образуются и какие функции выполняют?
10. На какой стадии эмбрионального развития находится яйцо зародыша птиц в период откладывания яйца?
11. Как осуществляется вторая фаза гаструляции у птиц, чем она отличается от таковой у пресмыкающихся?
12. Что называется первичной полоской? Опишите процесс ее образования.
13. Опишите процесс образования мезодермы у птиц.
14. Охарактеризуйте процесс образования провизорных органов у птиц.
15. Чем отличается процесс образования амниотической и серозной оболочек у птиц от таковых у пресмыкающихся? Назовите их функции.
16. Как у птиц образован барьер «кровь-воздух», и какие функции он выполняет?
17. Какие группы млекопитающих выделяют в связи с особенностями эмбрионального развития? Дайте им характеристику.
18. Охарактеризуйте процесс дробления у сумчатых млекопитающих.
19. Как происходит гаструляция у сумчатых млекопитающих?
20. Перечислите и охарактеризуйте особенности эмбрионального развития сумчатых млекопитающих.
21. Что представляет собой трофобласт сумчатых млекопитающих?
22. Охарактеризуйте полуплаценту сумчатых млекопитающих.
23. Опишите процесс образования хориона. Какие функции он выполняет?
24. Опишите процесс дробления яйцеклетки плацентарных млекопитающих.
25. Какие различия у плацентарных млекопитающих наблюдаются в строении бластомеров? Назовите и охарактеризуйте эти бластомеры. Какие функции они выполняют?
26. На какой стадии эмбрионального развития у плацентарных млекопитающих трофобласт вступает в контакт со слизистой оболочкой матки?
27. Что называется имплантацией? Когда она происходит у человека?
28. Опишите процесс гаструляции у плацентарных млекопитающих.
29. Какие различия зародышевых оболочек отмечаются у плацентарных млекопитающих по сравнению с пресмыкающимися, птицами, низшими млекопитающими?
30. Какие изменения у зародыша человека и приматов происходят в процессе дробления, начиная со стадии 12-клеточной морулы?
31. Что способствует ускоренному развитию зародыша человека?
32. Дайте характеристику зрелой бластоцисты зародыша человека.
33. Когда и как возникает амнион у зародыша человека и приматов?
34. Дайте характеристику первой фазы гаструляции у человека и приматов. Когда она происходит?
35. Что называется плазмодиотрофобластом, его структура и функции.
36. Охарактеризуйте синтициальный трофобласт.
37. Опишите процесс образования ворсинок у зародыша человека.
Если Вам понравилась эта лекция, то понравится и эта - Лекция 12.
38. Опишите вторую фазу гаструляции у зародыша человека.
39. Что представляет собой плацента у плацентарных млекопитающих, ее функции.
40. Какие процессы происходят у зародыша человека в сомитный период?
41. Опишите строение и функции пупочного канатика зародыша человека и приматов.
42. Перечислите и охарактеризуйте типы плацент млекопитающих животных.



















