
Механизмы переноса энергии и заряда в биомолекулярных системах
1.1 Механизмы переноса энергии и заряда в биомолекулярных системах
Наличие у биологически важных молекул коллективизированной системы π-электронов, способных принимать и отдавать энергию и заряд, позволяет им активно участвовать в разнообразных процессах жизнедеятельности. Здесь уже на квантовом уровне сталкиваемся с проявлением диалектического единства структуры и функции. Оно отображается так называемой “электронной схемой жизни”, иллюстрирующей значение коллективизированной системы p-электронов биомолекул в фотосинтезе и клеточном дыхании. При переносе подвижного электрона по цепи структурно связанных между собой макромолекул происходит преобразование солнечной энергии во все формы работы, совершаемой биологическими системами.
Для сложных органических молекул характерны внутримолекулярный и межмолекулярный переносы энергии и заряда. Так, каждая из двух или более автономных систем p-электронов молекулы может проявлять спектральную независимость и вместе с тем взаимодействовать с другими. Взаимодействие π-электронных систем внутри молекулы связано с миграцией энергии по ней, то есть с переносом энергии между разными функциональными группами этой молекулы. Например, в молекуле представляющей собой гибрид антрацена и нафталина, метиленовые группы “разобщают” коллективированные части молекулы (рис. 1.19). Однако эти группы не препятствуют переносу энергии внутри молекулы, о чём свидетельствует появление люминесценции антрацена при возбуждении молекулы в полосе поглощения нафталина.
Чрезвычайно важной для жизнедеятельности является способность биомолекул передавать энергию. Под миграцией энергии понимается безызлучательный обмен энергией между электронно-возбужденной молекулой – донором (D) и молекулой в основном состоянии – акцептором (А): D* + А ® D + А*.
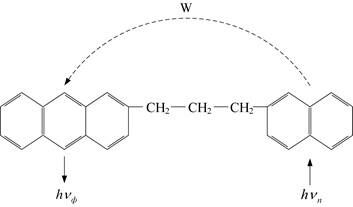
Рис. 1.19. Внутримолекулярный перенос энергии возбуждения.
Направление переноса показано пунктирной стрелкой
Миграция энергии – физический процесс, не сопровождающийся химическим изменением вещества. Она может происходить как между одинаковыми, так и между разными молекулами в направлении от более высокого к более низкому или одинаковому энергетическому уровню. Известно несколько механизмов миграции энергии: индуктивно-резонансный, обменно-резонансный, полупроводниковый и экситонный. Рассмотрим кратко эти механизмы.
Индуктивно-резонансная миграция энергии. Исходным фактом, который заставил принять концепцию индуктивно-резонансной миграции энергии, было явление сенсибилизированной флуоресценции красителей, то есть эффективное возбуждение флуоресценции молекул акцептора светом, поглощаемым молекулами донора. При определённых условиях к акцептору безызлучательным путем передавалась практически вся энергия донора через большие межмолекулярные расстояния (порядка 2 – 10 нм). Такая ситуация наблюдается только в тех случаях, когда спектр флуоресценции донора и спектр поглощения акцептора перекрываются. Эффективность миграции прямо пропорциональна площади перекрытая, то есть чем больше одинаковых для переходов D* ® D и А ® А* частот, тем выше вероятность резонанса. Однако этот процесс представляет собой более сложное явление, чем простая реабсорбция молекулой акцептора светового кванта, излучённого молекулой донора. При классическом рассмотрении можно считать, что электронные системы молекул D* и А представляют собой механические осцилляторы (наподобие маятников), способные колебаться с одинаковой частотой и взаимодействующие друг с другом. Когда молекула А оказывается поблизости от молекулы D*, появляется определённая вероятность того, что молекула D*, прежде чем испустить фотон, передаст акцептору свою энергию возбуждения.
Такому процессу присущи две важные особенности. Во-первых, перенос энергии происходит с очень высокой скоростью. Во-вторых, перенос энергии может происходить на довольно значительные расстояния. Например, эффективный перенос энергии в растворах наблюдается на расстояниях, превышающих 10 нм, причем для миграции энергии не требуется высокоупорядоченной организации молекул, участвующих в нём.
Обменно-резонансная или триплет-триплетная миграция энергии. Перенос энергии по триплетиым уровням молекул был впервые обнаружен А. Н. Теренииым и В. Л. Ермолаевым в 1952 г. При обменно-резонансной миграции энергия переносится с триплетного уровня молекулы D на триплетный уровень молекулы A(3D+1A®D+3A) не в результате индуктивно-резонансного взаимодействия молекул, а при прямом перекрывании «триплетных» электронных орбиталей (электронных облаков) за счет электростатических взаимодействий электронов донора и акцептора. Чем больше объём области перекрытия, тем вероятнее перенос, при котором партнеры обмениваются электронами: акцептор получает богатый энергией электрон донора, отдавая ему свой бедный энергией электрон. Отсюда возник и сам термин “обменно-резонансный” перенос.
Рекомендуемые материалы
Полупроводниковая миграция энергии (зонная проводимость). Как известно, в твердых телах энергетические уровни валентных электронов образуют энергетические зоны, внутри которых могут перемещаться электроны и дырки. Таким образом, в противоположность другим механизмам миграции энергии, полупроводниковый (зонный) перенос энергии связан с переносом зарядов, что делает возможным пространственное разделение окислителя (дырки) и восстановителя (электрона), а также длительное запасание энергии в ловушках электронов, в качестве которых могут выступать примеси.
С вопросом о миграции энергии и зарядов внутри молекул тесно связан вопрос о полупроводниковых свойствах биологически важных макромолекул. Так, p-электронное взаимодействие между основаниями в молекуле ДНК приводит к тому, что эту гигантскую молекулу можно рассматривать как одномерное твердое тело и изучать с позиций зонной теории. Проводимость кристаллов ДНК возрастает при облучении светом, что отвечает переходу электронов из заполненной валентной зоны в свободную зону. Не известно, правда, к какому типу полупроводников (п или р) можно отнести ДНК. Источником электронов, осуществляющих проведение тока, служат, по-видимому, примеси. Возможность движения электронов вдоль молекулы ДНК между нуклеотидными основаниями появляется благодаря перекрыванию p-электронных систем этих оснований.
Экситонная миграция. При образовании пары электрон - дырка в кристаллах, когда электроны переходят из валентной в свободную зону, возникают так называемые экситоны. Они образуются в тех случаях, если энергия возбуждения меньше, чем ширина запрещённой зоны. При этом электрон и дырка не могут независимо перемещаться в кристалле, а будучи связанными друг с другом, образуют электрически нейтральную квазичастицу – экситон.
Перемещаясь по кристаллу, экситон переносит энергию, но не переносит электрического заряда. Экситоны возникают обычно в молекулярных кристаллах, в которых внутримолекулярные связи значительно сильнее, чем связь молекул между собой. Такой тип миграции энергии часто наблюдается, например, в препаратах ДНК и белков, что подтверждает возможность необычайно быстрого и эффективного переноса энергии в высокоорганизованных молекулярных системах. Время экситонного переноса энергии составляет 10-14 с. Такая высокая скорость миграции позволяет полностью избежать потерь энергии возбуждения молекул. Экситонный перенос является одним из возможных механизмов переноса энергии при фотосинтезе.
Перенос заряда. Важным свойством сложных биомолекул, обладающих развитой системой двойных связей и делокализованной p-электронной системой, является их способность к межмолекулярному переносу заряда.
Рассмотрим кратко комплексы с переносом заряда. Комплекс – это пара взаимодействующих молекул, одна из которых отдает электроны (донор), а другая – принимает их (акцептор). Перекрывание молекулярных волновых функций, а с ним и вероятность переноса заряда сильно зависят от положения энергетических уровней донора и акцептора, в соответствии с чем различают несколько вариантов переноса.
Так, слабый перенос не может идти без поглощения квантов света. Условно здесь можно выделить два крайних случая. Первый из них: 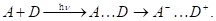
Молекулы А и D слабо взаимодействуют в основном состоянии. При возбуждении светом происходит фотоперенос электрона с заполненной молекулярной орбитали донора на свободную орбиталь акцептора, что проявляется в существовании у комплекса А ... D характеристической полосы поглощения, отсутствующей в спектрах изолированных молекул А и D (рис. 1.20, а). Такая полоса называется полосой переноса заряда (ПЗ). Её максимум приходится на область более длинных волн, чем максимум полосы поглощения донора. После облучения комплекса в полосе ПЗ донор и акцептор представляют собой своеобразную ионную пару, то есть в возбуждённом состоянии комплекс практически ионизируется под действием света.
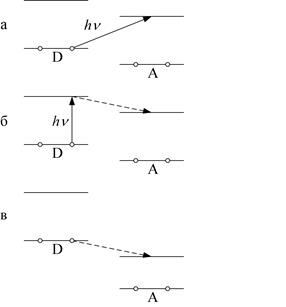
Рис. 1.20. Схема, иллюстрирующая образование комплекса с переносом заряда: а, б – слабый перенос, в – сильный перенос
Второй случай слабого переноса заряда можно изобразить так: 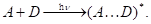
При электронном возбуждении одного из компонентов комплекса образуется его связь с другим компонентом. В этом процессе можно выделить два этапа. Вначале донор при поглощении света переходит в возбуждённое состояние: D D*. В возбуждённом состоянии D* взаимодействует с А и образует комплекс D* + А ® (А ... D)*. Схема уровней, между которыми происходит электронный перенос, показана на рис. 1.20, б.
D*. В возбуждённом состоянии D* взаимодействует с А и образует комплекс D* + А ® (А ... D)*. Схема уровней, между которыми происходит электронный перенос, показана на рис. 1.20, б.
Сильный, перенос электрона может происходить в темноте (рис. 3.20, в). Здесь при переходе электрона с орбитали донора на акцепторную орбиталь энергия выделяется, а не поглощается. Если она достаточна для преодоления электростатического притяжения электрона к донору, то электрон переносится на акцептор без освещения комплекса. Комплекс, образующийся при сильном переносе, способен диссоциировать в растворителе на два свободных радикала. Они интенсивно поглощают свет разных длин волн, в силу чего раствор приобретает окраску. Важно отметить, что перенос электронов по макромолекулам, сопровождающийся во многих случаях образованием свободных радикалов, имеющих высокую реакционную способность, является обязательным предшественником большинства биохимических реакций.
Способность к передаче энергии и заряда – очень важное свойство биомолекул, определяемое во многом особенностями их строения. Рассмотрим конкретный пример переноса энергии и заряда в биологической системе.
Перенос энергии и заряда при прямом действии ионизирующей радиации на белки и нуклеиновые кислоты. В результате одиночного взаимодействия кванта ионизирующего излучения с молекулой белка или нуклеиновой кислоты эти молекулы могут утратить свои функциональные свойства, если процесс идёт с поглощением энергии порядка 60 – 70 эВ. Между воздействием излучения на макромолекулу и её инактивацией происходит ряд последовательных физико-химических процессов. Кроме того, установлено, что инактивация белков и нуклеиновых кислот нередко связана с повреждением структуры макромолекулы, которое возникает не в любом, а определённом её участке. Селективность действия облучения объясняют миграцией поглощённой энергии по макромолекуле с локализацией её в определённом “слабом” звене, в котором и произойдут дальнейшие химические изменения. Таким “слабым” звеном может быть один из аминокислотных остатков, составляющих молекулу белка, или одно из нуклеотидных оснований, входящих в молекулу ДНК.
Рассмотрим некоторые закономерности ионизации и миграции энергии и заряда в молекулах белков. Эти молекулы имеют огромное количество химических связей и по структуре напоминают скорее “малые кристаллы”, чем молекулы. В полипептидной цепи белка атомы “соединены” ковалентными связями, обусловленными обменом электронов между двумя соседними атомами. В образовании таких связей участвуют как s-, так и p-электроны.
Предположим, что под действием гамма-кванта атом углерода, показанный справа на рис. 1.21, а, теряет электрон, который обеспечивал связь этого атома с соседними.
Если Вам понравилась эта лекция, то понравится и эта - Исследование личности потерпевшего.
В малой молекуле подобное событие привело бы к её распаду, тогда как в ионизированной макромолекуле атом, потерявший электрон, остается на своем месте (по крайней мере, в течение долей секунды), но несёт теперь положительный заряд.
Он эквивалентен дырке в полупроводнике. Поэтому электрон соседнего атома стремится перейти к атому с дыркой. Такое движение электрона соответствует перемещению дырки в противоположном направлении, например, к атому азота и далее (даже в боковую цепь – R – SH). Каждый такой “шаг” заряда занимает время, необходимое для одного электронного перехода (порядка 10-15 с). Миграция заряда по цепи равносильна переносу “избыточной” энергии возбуждения. Благодаря миграции энергии и заряда дырка достигает любого доступного места цепи в течение примерно 10-12 с. Мигрирующий заряд, если на его пути встречается “слабое” звено, локализуется там, прекращая дальнейшее перемещение. Локализация положительного заряда у определённого атома, например, у атома серы в группе – SH, приводит к разрыву в этом месте химической связи и образованию неспаренного электрона в атоме серы и атоме водорода. Возникает сложный биологический радикал белка, включающего серу, и малый радикал водорода. Неспаренные электроны могут образоваться и в других атомах, например, в атомах азота и углерода. Белковая молекула в свободнорадикальном состоянии обладает повышенной реакционной способностью. Поэтому такие белки-радикалы мгновенно вступают в разнообразные химические реакции (например, реакции присоединения). Это в конечном итоге приводит к изменению структуры белковой молекулы и потере ею биологической функции.
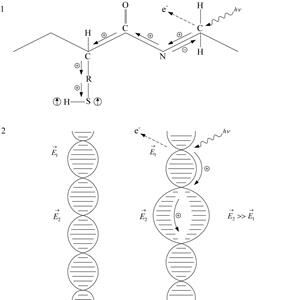
Рис. 1.21. Действие ионизирующего излучения на белки (1) и нуклеиновые кислоты (2). Направление миграции дырок в молекуле указано стрелками. Стрелка в кружке обозначает неспаренный электрон.  – локальные напряжённости электрического поля
– локальные напряжённости электрического поля
Аналогичный механизм ионизации и миграции дырки присущ нуклеиновым кислотам при действии на них ионизирующего излучения. Так, дырка, возникшая при поглощении энергии ионизирующей радиации в определённом месте молекулы ДНК и создавшая там электрическое поле напряжённостью Е1, движется вдоль оси молекулы и локализуется у пары азотистых оснований (нуклеотидов), где напряжённость поля Е2 выше, чем Е1. Если Е2 становится достаточной для разрыва химических связей и разделения пары оснований из-за сильной поляризации p-орбитали, то следствием локализации дырки будет расплетение ДНК (рис. 1.21, б) и утрата биологической функции.
Миграции заряда по молекулам белков и нуклеиновых кислот способствует перекрывание π-орбиталей в них. Потенциальные энергетические барьеры, встречающиеся на пути движения зарядов, преодолеваются, по-видимому, по механизму туннельного эффекта.



















