
Самоорганизация белка
31. Самоорганизация белка. Этапы самоорганизации белка. Термодинамическая оценка процесса самоорганизации. Особенности процесса самоорганизации белка в условиях in vivo. Парадокс Левинталя, его сущность и разрешение.
Самоорганизация – это путь к глобальному минимуму свободной энергии. Этот путь единственно возможный в данных условиях для данной молекулы. Для белков процесс самоорганизации заключается в образовании пространственной структуры.
В процессе ренатурации белка разные стадии представляют собой фазовые переходы разного рода. Для малых белков процесс можно считать одностадийным. Этот процесс происходит как ФП 1 рода.
Для крупных белков этот процесс многостадийный, и разные его стадии происходят как ФП разного рода.
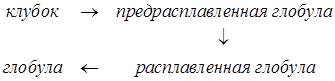
Ранние этапы самосборки различаются в зависимости от типа вторичной структуры.
Образование α-спирали происходит как нефазовый переход, так как оба фазовых состояния одномерны и не происходит изменения границы фаз.
Если Вам понравилась эта лекция, то понравится и эта - Содержание.
Образование β-листов происходит как ФП 1 рода, площадь контакта цепи с листом зависит от размера контактирующих элементов. Процесс образования β-структуры происходит значительно дольше.
Процесс образования нативной структуры из расплавленной глобулы происходит как ФП 1 рода. В процессе этого перехода происходит уменьшение энтропии системы, компенсированное падением свободной энергии. Это обеспечивается наличием большого числа слабых связей с низкой энергией.
К ФП 2 рода могут относиться процессы образования и разрушения доменных структур, которые часто сопровождаются ростом теплоёмкости системы.
Парадокс Левинталя заключается в том, что каждый АК остаток имеет порядка 10 возможных конформаций, цепь из 100 АК остатков будет иметь 10100 возможных конформаций. При этом, белок должен менять свою конформацию в сторону наименьшей свободной энергии и "почувствовать" стабильное состояние. При этом перебор возможных состояний даже короткой цепи в 100 АК потребовал бы 1080 лет при скорости перехода между конформациями порядка 10-13 секунд. Но при этом процесс образования нативной структуры происходит за время порядка 1 секунды.
Парадокс разрешается при учёте нуклеационного механизма сворачивания. Пространственная организация начинается в самом начале синтеза полипептидной цепи с образования ядра сворачивания. Ядро сворачивания образуется изначально из АК имеющих небольшое число разрешённых конформаций. В процессе синтеза цепи новые АК контактируют с ядром сворачивания, которое ограничивает число разрешённых конформаций. В процессе синтеза всё новые АК вовлекаются в ядро сворачивания, увеличивая его поверхность, скорость падения энергии системы возрастает. В этом заключается котрансляционный механизм сворачивания. Образование зародыша сворачивания соответствует локальному энергетическому минимуму, а далее система стремится к глобальному энергетическому минимуму – к нативной структуре.
Также в клетке могут присутствовать ферменты, катализирующие отдельные этапы самосборки – фолдазы, а также шаперолы, белки теплового шока, способные стабилизировать структуру синтезируемого белка, формируя с ним нековалентные связи.


















