
Динамика острого воспалительного процесса
3. Динамика острого воспалительного процесса
3.1. Этиология воспаления
Любое внешнее (экзогенное) или внутреннее (эндогенное) воздействие на ткани организма, превышающее их адаптационные возможности, способно вызвать повреждение клеток и внеклеточных структур и, как следствие, привести к воспалению. Экзогенные повреждающие факторы могут иметь механическую, физическую, химическую или биологическую природу (микробы, вирусы, грибы, простейшие, черви, насекомые). Эндогенные повреждающие факторы образуются непосредственно в самом организме в результате определенной патологии. Например, воспалительный процесс может возникнуть в миокарде как результат некроза кардиомиоцитов после перенесенного инфаркта миокарда. В этом и других подобных случаях воспаление будет носить название «асептического воспаления».
3.2. Патогенез и стадии острого воспалительного процесса
В приведенном выше определении воспаления было указано, что воспаление характеризуется явлениями альтерации, экссудации и пролиферации. Именно эти три стадии воспаления, накладывающиеся друг на друга и сменяющие одна другую, принято считать основными этапами развития острого воспалительного процесса. Следует учесть, что пусковым моментом для начала каждой из этих трех стадий является повреждение ткани экзогенным или эндогенным повреждающим фактором. Так, например, практически сразу после альтерации ткани на периферии воспалительного очага можно наблюдать явления пролиферации, а явления экссудации начинают проявляться в самом начале вторичной альтерации. Однако каждая из стадий достигает максимума развития в различное время и именно это обстоятельство позволяет говорить об определенной сменяемости стадий воспалительного процесса.
В дальнейшем, разбирая особенности патогенеза воспаления, мы будем соотносить и привязывать отдельные звенья патогенеза к основным стадиям воспаления, а именно – к альтерации, экссудации и пролиферации. Следует указать, что этому правилу следуют и большинство патологов и клиницистов при описании воспалительного процесса. Существуют и другие принципы изложения материала по проблеме патогенеза воспаления. Так, например, А.М.Чернух выделяет в развитии воспаления пять стадий, основываясь на особенностях реакций микроциркуляторного русла в очаге воспаления. По нашему мнению этот принцип в определенной степени сужает подход к описанию патогенеза воспаления, так как, явления альтерации, экссудации и пролиферации определяются чрезвычайно большим количеством разнообразных действующих факторов, в число которых, без сомнения, входит и реакция микроциркуляторного русла.
3.2.1. Первичная и вторичная альтерация
В стадии альтерации выделяют две взаимосвязанные фазы: фаза первичной альтерации и фаза вторичной альтерации. Первичная альтерация возникает в результате непосредственного действия на ткань повреждающего фактора. При этом часть клеток разрушается самим повреждающим фактором, а часть повреждается в большей или меньшей степени за счет развившейся непосредственно после повреждения локальной гипоксией, которая обусловлена ишемией ткани (активный нейрогенный спазм, как поврежденных сосудов, так и сосудов, находящихся в непосредственной близости от участка повреждения). Гипоксический некробиоз клеток, прежде всего, сказывается на состоянии их мембран: плазматической мембраны и внутриклеточных мембран. Повреждение плазматической мембраны приводит к нарушению разделения ионов между протоплазмой клеток и внеклеточной средой. В результате в клетках происходит накопление ионов Na+, а вслед за ними - и воды, и ионов Са++. Повышенная гидратация клеток способна в конечном итоге вызвать их осмотический взрыв, а повышенное поступление ионов Са++ в их протоплазму активирует мембранные фосфолипазы и запускает процесс образования продуцентов арахидоновой кислоты – значимых медиаторов воспаления1/. С другой стороны, повреждение внутриклеточных мембран может так же привести к необратимому повреждению клеток. Особенно опасно повреждение мембран лизосом и митохондрий. Гидролитические ферменты, содержащиеся в лизосомах, выходят в цитоплазму и вызывают разрушение веществ, входящих в состав
________________________________
Рекомендуемые материалы
1/ Описание медиаторов воспаления и, в частности, медиаторов арахидонового каскада, смотри в разделе «3.2.1.1. Медиаторы воспаления»
цитоплазмы и клеточных органелл. Клетки лизируются («самоперевариваются») и фермен-
ты лизосом (протеазы, липазы, гликозидазы, фосфатазы) выходят в межклеточную среду, нанося повреждение и другим близлежащим клеткам. Повреждение митохондрий резко нарушает энергообмен клеток, тормозится процесс окислительного фосфорилирования, снижается синтез АТФ. Энергетическое голодание клеток так же способно привести к их гибели. Кроме того, повреждение митохондрий нарушает утилизацию ими жирных кислот и, при определенных условиях, стимулировать образование в митохондриях активных кислородных радикалов – активных форм кислорода (АФК), повреждающее воздействие на клетки которых хорошо известно.
Гипоксия и, следующий за нею, гипоксический некробиоз клеток повреждают не только такие клеточные органеллы как лизосомы и митохондрии. Разрушается цитоскелет клеток, гладкий и шероховатый эндоплазматический ретикулум, другие клеточные органеллы.
Подведем некоторые итоги. Первичная альтерация – это повреждение и гибель клеток в результате непосредственного воздействия на ткань любого повреждающего фактора. В результате этого процесса в межклеточную среду выходят или образуются в ней многочисленные биологически активные вещества, способные сами по себе, уже в условиях прекращения действия повреждающего фактора, продолжать разрушать клетки и другие межклеточные структуры. Иначе говоря, вслед за первичной альтерацией начинает развиваться вторая фаза альтерации – вторичная альтерация.
Большинство биологически активных веществ как разрушающих клетки и межклеточные структуры, так и участвующих в регуляции воспалительного процесса в целом, выделяются, активируются и начинают осуществлять свое действие именно во время развития вторичной альтерации. Эти биологически активные вещества получили название медиаторов воспаления. Рассмотрим основные медиаторы воспаления.
3.2.1.1. Медиаторы воспаления
Существует достаточно много классификаций медиаторов воспаления. В качестве примера приведем следующие классификации медиаторов: по их химическому строению, по скорости включения в процесс воспаления, по принципу действия – прямому или опосредованному (в первом случае медиатор участвует в регуляции воспалительного процесса как непосредственно действующее вещество, во втором – является источником высвобождения или образования некоего другого медиатора). Однако наиболее традиционной и часто применимой патологами и клиницистами является разделение медиаторов на две группы: медиаторы клеточные и медиаторы гуморальные (или плазменные).
Медиаторы, относящиеся к первой группе, появляются в очаге воспаления или при разрушении соответствующих клеток, или секретируются (выделяются) ими в результате воздействия на них повреждающего фактора или определенных биологически активных веществ. Медиаторы второй группы образуются в плазме крови (поэтому они и называются «плазменными») в ходе некоторых биохимических процессов, которые инициируются рядом факторов воспаления.
Характерной чертой всех клеточных медиаторов является то, что они образуются практически сразу же после воздействия на ткань повреждающих факторов воспаления и действуют локально, то есть непосредственно в зоне контакта повреждающего фактора с тканью.
Образование гуморальных (плазменных) медиаторов происходит через определенный промежуток времени после воздействия повреждающего фактора (каждой биохимической реакции требуется время для ее завершения). Кроме того, гуморальные медиаторы, за счет их образования в плазме крови, обладают более системным действием, чем клеточные медиаторы.
Рассмотрим основные медиаторы, имеющие клеточное происхождение.
Гистамин. Этот медиатор воспаления синтезируется и гранулируется в тучных клетках (лаброцитах) и в базофилах. Гистамин синтезируется из гистидина под влиянием фермента гистидиндекарбоксилазы. По химическому строению гистамин относится к группе биогенных аминов.
Описано существование гистамина в трех формах: связанный, лабильный и свободный. Связанный гистамин может освобождаться только при разрушении клетки-носителя, лабильный высвобождается при действии на тучные клетки либераторов гистамина (например, лизосомальных ферментов), вызывающих их дегрануляцию. В дегрануляции лаброцитов и движении гранул к мембране клетки принимают участие ионы Са++, которые активируют внутриклеточные микромиофиламенты, с помощью которых гранулы доставляются к плазматической мембране. Свободный гистамин содержится в органах и тканях лишь в незначительном количестве.
Так как тучные клетки располагаются в непосредственной близости от микрососудов, эффекты гистамина сказываются, прежде всего, именно на них, причем от времени действия повреждающего фактора на ткань и до начала секреции гистамина проходит всего несколько секунд. Расширение сосудов осуществляется за счет действия гистамина на Н1 и Н2 гистаминорецепторы (в основном – через Н1 гистаминорецепторы). В стадию артериальной гиперемии гистамин обеспечивает повышенный приток крови к очагу воспаления за счет раскрытия прекапиллярных сфинктеров, расширения капилляров и, особенно, посткапиллярных венул. Второе важное направление действия гистамина – повышение проницаемости микрососудов за счет увеличения подвижности эндотелиальных клеток, их округления и, вследствие этого, появления промежутков между ними. Помимо этого гистамин стимулирует фагоцитоз, усиливает хемотаксис фагоцитов и митогенез лимфоцитов. Следует указать, что действие гистамина весьма краткосрочно, так как он очень быстро разрушается соответствующими ферментами, и в дальнейшем сосудистые реакции в очаге воспаления поддерживаются другими медиаторами.
Серотонин (5-гидрокситриптамин). Этот медиатор воспаления образуется из триптофана путем декарбоксилирования. По химическому строению относится к группе биогенных аминов. У человека серотонин содержится в тромбоцитах и тучных клетках кожи. Высвобождение серотонина из клеток-носителей происходит или при их разрушении, или под влиянием некоторых биологически активных веществ (например, тромбина, АДФ, фактора активации тромбоцитов - ФАТ). В очаге воспаления серотонин обеспечивает расширение артериол и повышает проницаемость микрососудов. Кроме того, серотонин способствует спазму венул и тромбообразованию в этих посткапиллярных сосудах, что и обеспечивает развитие венозной гиперемии при воспалении.
Ферменты лизосом. Лизосомы тканевых клеток, а также гранулоцитов, тучных клеток и базофилов играют важную роль в развитии воспалительного процесса. При разрушении тканевых клеток под влиянием повреждающих факторов воспаления, а также в результате фагоцитоза и клеточного киллинга, ферменты лизосом (протеазы, липазы, фосфатазы, гликозидазы) выходят в межклеточную среду, где становятся одним из основных факторов вторичной альтерации и экссудации, так как ферменты, содержащиеся в этих органеллах, способны дегранулировать тучные клетки, активировать кининовую систему плазмы крови и, за счет действия фосфолипаз, запускать каскад образования ряда биологически активных веществ, синтезирующихся из фосфолипидов клеточных мембран. Именно поэтому лизосомы принято называть «пусковыми площадками воспаления». Кроме того, лизосомные ферменты являются инициаторами образования таких мощных факторов вторичной альтерации как активные кислородные радикалы (АКР).
Велико значение ферментов лизосом и в процессе повышения проницаемости микрососудов в очаге воспаления. С одной стороны, они влияют на этот процесс косвенно (дегрануляция тучных клеток, активация кининовой системы и арахидонового каскада), а с другой – непосредственно, за счет разрушения (перфорации) базальной мембраны микрососудов (фермент лизосом – химаза способна разрушать хондроитинсульфаты, входящие в состав соединительно-тканного вещества базальной мембраны). Следует иметь в виду, что большинство лизосомальных ферментов наиболее активны в кислой среде, которая и характерна для очага острого воспаления.
Активные кислородные радикалы (АКР) принимают активное участие в повреждении клеток и внеклеточных структур в процессе вторичной альтерации. По современным представлениям свободный радикал (в том числе – и АКР) – это атом или молекула, имеющие неспаренный электрон на внешней орбите. В частности, этими свойствами обладают супероксидный анион – О2+ - и синклетный кислород – 1О2. Повышенная окислительная способность придает свободным радикалам особую химическую агрессивность и позволяет им не только активно вступать в реакции с клеточными структурами (например, при перекисном окислении липидов мембран клетки), но и превращать молекулы клеточных структур в новые свободные радикалы. Так возникает своеобразная «цепная реакция» генерирования свободных радикалов в клетке. Возникая в очаге воспаления во время и благодаря процессам вторичной альтерации, АКР разрушают клетки не избирательно, но действуют даже и на те клеточные образования, которые не имеют достаточной антиоксидантной защиты и находятся на некотором отдалении от зоны воспаления.
Производные арахидоновой кислоты (эйкозаноиды). В очаге воспаления производные арахидоновой кислоты (эйкозаноиды) синтезируются под влиянием фосфолипазы А2, лизирующей фосфолипиды мембран клеток и становящейся особенно активной при избытке ионов Са++, которые в больших количествах освобождаются из погибших клеток. Чаще всего сигналом для активации фосфолипазы А2 служит воздействие на клетки ферментов лизосом. Образовавшаяся из фосфолипидов арахидоновая кислота под влиянием двух ферментов липооксигеназы и циклооксигеназы расщепляется, образуя две группы биологически активных веществ: простагландинов и лейкотриенов.
Функции простагландинов весьма обширны. Так, простагландины группы Е вызывают расширение сосудов, потенцируя действие гистамина и серотонина, обладают хемотаксическим действием по отношению к поли - и мононуклеарам крови. На стадии же пролиферации простагландины этой группы усиливают синтез коллагена фибробластами. Участвуя в сложных взаимоотношениях с рядом цитокинов, простагландины этой группы возбуждают лихорадочную реакцию.
Другие производные арахидоновой кислоты – лейкотриены (фракции В4, С4, D4, Е4). При развитии воспаления лейкотриены повышают сосудистую проницаемость, увеличивают активность клеток – естественных киллеров. Побочным и весьма неприятным эффектом действия лейкотриенов является их влияние на гладкую мускулатуру внутренних органов.
В частности, лейкотриены вызывают бронхоспазм.
К гуморальным (плазменным) медиаторам воспаления относятся следующие биологически активные вещества:
Плазменная система кининов. В целом, образование и метаболизм кининов в очаге воспаления можно представить следующим образом. При повреждении стенок микрососудов в процессе развития вторичной альтерации, активируется XII фактор системы свертывания крови (фактор Хагемана). Функции фактора Хагемана весьма разнообразны. Во-первых, он является одним из необходимых участников процесса свертывания крови, то есть образования тромбов на стенках микрососудов. Во-вторых, при его участии активируется фермент плазмин, благодаря которому запускается система фибринолиза. И, наконец, в-третьих, при его содействии протеолитические ферменты, входящие в группу калликреинов (калликреин-1, калликреин-2) и находящиеся в неактивном состоянии в форме прекалликреинов (калликреиногенов), активируются и, далее, обеспечивают превращение кининогена (a2-глобулина плазмы крови) в активные кинины (брадикинин и каллидин).
Активные кинины выполняют функцию вазодилатации и повышения проницаемости сосудов и довольно быстро разрушаются под действием разнообразных кининаз. Однако существует и своеобразный порочный круг: кинины активируют фактор Хагемана, а он, в свою очередь, участвует в переводе кининов в активную форму. Кроме того, есть данные о том, что в процессе активации кининов участвует гистамин, протеазы распадающихся при воспалении клеток, катионные белки лейкоцитов и некоторые другие вещества, образующиеся в очаге воспаления.
Система комплемента. Система комплемента – это часть иммунной системы, осуществляющая неспецифическую защиту от бактерий и других, вредных для организма антигенов. Она состоит более чем из 20 различных белков — факторов (компонентов) комплемента, находящихся в плазме крови и составляющих около 4% от всех белков плазмы.
Система комплемента участвует в регуляции воспалительного процесса следующими тремя способами:
- хемотаксис: факторы комплемента могут привлекать иммунные клетки, которые фагоцитируют бактерии;
- лизис: компоненты комплемента воздействуют на бактериальные мембраны и лизируют (растворяют) бактерии;
- опсонизация: компоненты комплемента, воздействуя на бактериальные клетки, облегчают их фагоцитирование.
Компоненты комплемента с С1 по С9 (С – от английского слова. complement) участвуют в так называемом «классическом пути» активации комплемента. Факторы В и D активируют «альтернативный путь» (Рис. 1). Другие компоненты системы комплемента обладают регуляторными функциями.
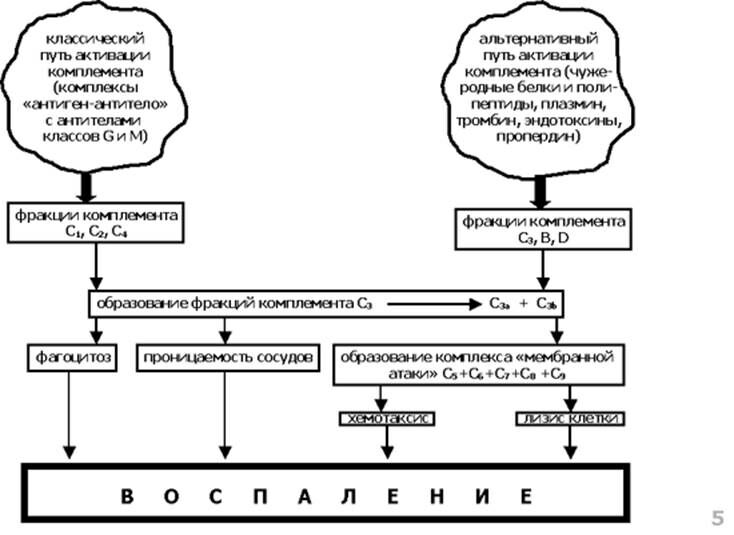
Рис. 1. Механизм действия системы комплемента в очаге воспаления
Классический путь начинается связыванием компонента C1 с несколькими молекулами иммуноглобулинов (IgG или с IgM) на поверхности бактерии. На альтернативном пути происходит связывание фактора В, например, с бактериальными липополисахаридами. Оба пути ведут к разделению компонента С3 комплемента на два фрагмента, обладающих различными функциями. Меньший фрагмент С3а принимает участие в воспалительном процессе, обуславливая хемотаксическое привлечение лейкоцитов к очагу воспаления. Более крупный фрагмент С3b «запускает» цепь реакций, приводящих к образованию так называемого мембраноатакующего комплекса комплемента.
Мембраноатакующий комплекс — это ионный канал в плазматической мембране бактериальной клетки, в образовании которого участвуют компоненты системы комплемента С3b, С5b, С6, С7, С8 и С9. В результате этой «атаки» меняются осмотические параметры бактериальной клетки, в неё входит большое количество воды, в результате чего возникает «осмотический взрыв» бактериальной клетки, и она гибнет.
Система комплемента контролируется её ингибиторами в плазме крови, которые блокируют избыточную активность этой системы.
Цитокины. Помимо указанных выше медиаторов воспаления, значительную роль в развитии и регуляции воспалительной реакции играют цитокины – низкомолекулярные 6елки (полипептиды или гликополипептиды с молекулярным весом 5-30 кДа), лишенные антигенной специфичности и являющиеся посредниками межклеточных взаимоотношений при воспалении, формировании иммунного ответа организма, гемопоэзе и ряде других межклеточных и межсистемных взаимодействий. Цитокины нельзя отнести и к клеточным медиаторам воспаления, и к гуморальным. Они занимают особую «нишу» в регуляции воспаления.
Несмотря на весьма значительные функциональные различия цитокинов, их о6ъединяют несколько важных признаков. Так, для цитокинов характерна функциональная взаимозаменяемость. Кроме того, при участии в регуляционных процессах цитокины способны к синергuзму uлu к антагонизму. Некоторые цитокины могут индуцировать синтез других цитокинов, активируя для этого соответствующие клетки иммунной системы. Все цитокины обладают коротким периодом действия.
Существует несколько классификаций цитокинов. Однако наиболее обоснованной нам видится следующая:
- Интерлейкuны (IL). В настоящее время описано 18 видов (от IL-1 до IL-18) интерлейкинов.
- Колонuестuмулuрующuе факторы (CSFs). Эти цитокины являются факторами роста гемопоэза (лимфопоэза, монопоэза, гранулопоэза).
- Интерфероны (IFNs). Интерфероны активируют естественные клетки-киллеры, инги6ируют репродукцию вирусов и могут участвовать в генерации ряда других цитокинов, активируя соответствующие клетки иммунной системы.
- Факторы некроза опухолей (TNFs). Из многочисленных функций этих цитокинов следует выделить их способность противостоять инфекционному началу и о6ладание противоопухолевой активностью.
- Хемокuны. Основной функцией многих цитокинов, входящих в эту группу, является стимуляция хемотаксиса практически всех клеток иммунной системы.
Как и каждая классификация, приведенное выше разделение цитокинов на некие функционально однородные группы - весьма условно. Так, интерлейкины 3, 7 и 11, помимо выполнения других функций, участвуют в гемопоэзе, а интерлейкины 8 и 16 о6еспечивают хемотаксис нейтрофилов, Т-лимфоцитов, 6азофилов и эозинофилов.
Оценивая роль цитокинов в воспалительном процессе, следует указать на одну важную особенность этих межклеточных посредников. В настоящее время можно выделить две группы цитокинов, одна из которых обладает провоспалuтельным действuем, а другая - протuвовоспалuтельным.
Провоспалuтельным действuем обладают интерлейкины 1, 6, 8, 12, 17, 18, гамма-интерферон, факторы некроза опухолей альфа и бета, фактор гемопоэза GM-CSF.
Протuвовоспалuтельным действuем обладают инги6итор интерлейкина 1 - IL-1ra, интерлейкин 10, трансформирующий фактор роста - бета (TGFb), интерфероны альфа, бета и дельта.
Провоспалительные эффекты цитокинов связаны с их возможностями активировать клетки иммунной системы, способствовать их дифференцировке, стимулировать выработку иммуноглобулинов, обеспечивать адгезию и хемотаксис фагоцитов. С другой стороны, чрезмерная активность провоспалительных цитокинов, недостаточное действие их инги6иторов и антагонистов может приводить к значительной деструкции тканей, росту альтерации. Не6лагоприятна роль провоспалительных цитокинов и при развитии хронического воспаления.
Протuвовоспалительные цитокины ограничивают развитие воспалительного процесса. Так, IL-1ra инги6ирует действие интерлейкина 1; интерлейкин 10 инактивирует макрофаги; TGF-b подавляет активность клеток - естественных киллеров, снижает пролиферацию В- и Т-лимфоцитов, инги6ирует макрофагальный киллинг внутриклеточных паразитов; интерфероны альфа, 6ета и дельта, в противоположность гамма-интерферону, обладают противовоспалительной активностью.
При не осложненном течении воспалительного процесса про- и противовоспалительные цитокины как 6ы уравновешивают друг друга, действуя на 6лаго организма. Смещение же этого 6аланса в лю6ую сторону может привести только к разрастанию патологии: мощной альтерации тканей или же к длительному, вяло текущему хроническому воспалению, зачастую осложняющемуся аутоиммунными процессами.
В заключение следует указать, что ряд цитокинов (интерлейкины 1 и 6, фактор некроза опухолей альфа, гамма-интерферон) являются эндогенными пирогенами, т.е. веществами, вызывающими лихорадочную реакцию, как правило, сопровождающую воспаление.
Белки острой фазы воспаления. В патофизиологии и клинике принято называть реакцию организма, следующую непосредственно вслед за альтерацией и направленную на восстановление гомеостаза организма, «реакцией острой фазы воспаления», а ряд биологически активных веществ, вырабатывающихся в этот период, «белками острой фазы».
К числу факторов, способных индуцировать реакцию острой фазы, относятся бактериальные и, в меньшей степени, вирусные инфекции, травмы, ожоги, злокачественные новообразования, тканевые инфаркты, воспалительные состояния. Реакция острой фазы включает такие клинические признаки и симптомы как сонливость, анорексию, изменения синтеза белков плазмы и синтеза некоторых гормонов.
Однако, прежде всего, острофазовая реакция характеризуется изменениями концентрации в плазме крови некоторых секреторных белков, вырабатываемых гепатоцитами при действии на печень цитокинов и некоторых гормонов. При этом основными индуктороми синтеза острофазных белков считается интерлейкин 1, интерлейкин 6, интерлейкин 11, гамма-интерферон, фактор некроза опухолей. Белки острой фазы, число которых весьма велико (более 20) разделяются на две группы: позитивные острофазные белки (концентрация их в плазме крови в процессе развития острофазовой реакции увеличивается в сотни и тысячи раз) и негативные острофазовые белки (их концентрация в плазме крови в процессе развития острофазовой реакции не изменяется, или даже уменьшается по сравнению с нормой).
Одной из основных функций острофазовых белков является модуляция воспалительной реакции и регенерации тканей. К «главным» белкам острой фазы относят С-реактивный белок и сывороточный амилоид А, концентрация которых в плазме крови после повреждения (воспаление, травма) в течение 6 – 8 часов возрастает в 100 – 1000 раз. С-реактивный белок способен связывать широкий спектр лигандов-компонентов микроорганизмов, токсинов, частиц поврежденных тканей, препятствуя тем самым их распространению. Кроме того, продукты такого взаимодействия активируют комплемент по классическому пути, стимулируя процессы фагоцитоза и элиминации вредных продуктов. С-реактивный белок может взаимодействовать с Т-лимфоцитами, фагоцитами и тромбоцитами, регулируя их функции в процессе воспаления. По видимому, белки острой фазы и некоторые пирогенные цитокины связаны между собой регуляционной обратной связью Так, С-реактивный белок вызывает увеличение синтеза фактора некроза опухолей макрофагами.
Следует указать, что С-реактивный белок обладает выраженными противовоспалительными функциями. В частности, он способен снижать высвобождение провоспалительных цитокинов из моноцитов, блокировать высвобождение фактора некроза опухолей из лейкоцитов ингибировать выработку тромбина и предохранять целостность сосудистого эпителия от альтерирующего воздействия на него медиаторов и цитокинов.
Сывороточный амилоид А способен усиливать адгезивность и хемотаксис фагоцитарных клеток и лимфоцитов. Кроме того, сывороточный амилоид А участвует в окислении липопротеинов низкой плотности и, тем самым, обладает антиатерогенным действием.
Таким образом, белки острой фазы при развитии местного острого воспаления регулируют его развитие, не допуская чрезмерной альтерации тканей и не допускают генерализации воспалительного процесса за счет своей противовоспалительной активности.
3.2.2. Экссудация
Развитие стадии экссудации (от exudare – «выпотевать») начинается в момент перехода первичной альтерации во вторичную и достигает максимума одновременно с пиком вторичной альтерации. Описывая стадию экссудации необходимо обратить внимание на два важных аспекта в развитии этого явления. Во-первых, в процессе экссудации жидкая часть крови, благодаря расширению и повышению проницаемости микрососудов, а также в силу изменения физико-химических характеристик крови и межклеточной жидкости, покидает сосудистое русло, образовывая воспалительный отек («плазматическая экссудация»). Во-вторых, кровеносное русло покидают и передвигаются к очагу воспаления такие форменные элементы крови как лейкоциты («клеточная инфильтрация»). В основе и плазматической экссудации, и клеточной инфильтрации лежат определенные патологические механизмы и закономерности.
С момента начала стадии экссудации проницаемость микрососудов претерпевает определенные изменения (Рис.2).

Рис. 2. Фазы повышения проницаемости сосудов при воспалении (по S.L.Robbins)
Пояснения в тексте.
Повышение проницаемости сосудов начинается на пике артериальной гиперемии и продолжается, постепенно затухая, вплоть до начала завершающей стадии воспаления, когда в действие вступают механизмы пролиферации и регенерации.
Повышение проницаемости сосудов проходит в несколько фаз.
Первая, или ранняя транзиторная фаза (кривая «А» на Рис.2) в основном обусловлена действием гистамина и серотонина и захватывает посткапиллярные венулы диаметром не более 100 мкм. Первая фаза длится не более нескольких минут.
На ее фоне развивается немедленная, длительная фаза повышения проницаемости (кривая «Б» на Рис.2). Она захватывает капилляры и вызвана повреждающим действием на микрососуды факторов, инициирующих воспалительную реакцию (некроз эндотелиальных клеток на уровне артериол небольшого диаметра).
Третья, отсроченная, стойкая фаза (кривая «В» на Рис.2) повышения проницаемости развивается через часы или даже сутки после начала воспаления. В ее основе лежит действие на сосуды простагландинов, лейкотриенов и других производных арахидоновой кислоты.
Выходу жидкой части крови за пределы микрососудистого русла способствует расширение капилляров и венул. Рассмотрим подробнее изменение тонуса микрососудов и их проницаемости в процессе развития воспаления.
3.2.2.1. Сосудистая реакция при воспалении
Выше уже было указано, что развитие первичной альтерации сопровождается спазмом (ишемией) артериол в зоне повреждения ткани. Ишемия вызывается быстрой (в течение одной – двух секунд) реакцией симпатической нервной системы на повреждение, выделением катехоламинов (норадреналина) и спастических агентов, продуцируемых эндотелием поврежденных микрососудов. Продолжительность ишемии не велика, так как норадреналин достаточно быстро разрушается ферментом моноамиоксидазой, которая образуется в очаге повреждения. Ишемию сменяет следующая фаза сосудистой реакции – расширение артериол, капилляров и венул. Развивается нейротоническая артериальная гиперемия, порождаемая парасимпатическими влияниями на тонус сосудов, осуществляемыми по принципу аксон-рефлекса. Нейротоническая артериальная гиперемия сменяется нейропаралитической артериальной гиперемией, в основе которой лежит утрата гладкомышечными элементами микрососудов способности реагировать на спастические влияния симпатической нервной системы. Именно поэтому, эту фазу сосудистой реакции еще называют «миопаралетической». Помимо влияния на тонус сосудов, повреждение ткани активирует и систему свертывания крови. Особенно активно этот процесс происходит в венулах – том участке микрососудистого русла, где кровоток наиболее замедлен, а стенки сосудов наиболее ранимы (более подробно описание процесса свертывания крови, возникающего в результате повреждения ткани, можно найти в разделе «Патология гемостаза. Тромбозы»). Тромбоз венул сначала замедляет, а затем и останавливает кровоток. В результате развивается сначала смешанная (артерио-венозная), а затем и венозная гиперемия, сменяемая стазом кровотока.
Одновременно с изменением тонуса микрососудов происходит и изменение их проницаемости. В этом процессе ведущую роль играют уже описанные выше медиаторы воспаления: гистамин, серотонин, ферменты лизосом, простагландины, активные кинины, факторы комплемента. В зависимости от степени проницаемости сосудов в очаге воспаления может образовываться транссудат (содержание белка до 2%), или экссудат (содержание белка до 6%). При значительных повреждениях сосудистой стенки кровеносное русло могут пассивно покидать и такие форменные элементы крови как эритроциты.
Характеристики жидкости, скапливающийся в воспалительном очаге, позволяют определить вид воспалительного процесса. В результате воспаление можно называть:
1. Серозным воспалением – в отеке содержится транссудат или экссудат, содержащий белок и не содержащий форменных элементов крови.
2. Фибринозным воспалением, когда в отечной жидкости содержится значительное количество фибрина, выпадающего в осадок на воспаленных тканях в виде нитей и пленок.
3. Гнойным воспалением, при котором в отечной жидкости содержится большое количество лейкоцитов, в основном погибших.
4. Геморрагическим воспалением - с отечной жидкостью, содержащей эритроциты (кровь в экссудате).
5. Ихорозным воспалением, когда в отечной жидкости поселяется гнилостная микрофло-
ра.
Значительную роль в переходе жидкой части крови через сосудистую стенку играют физико-химические изменения в очаге воспаления.
3.2.2.2. Физико-химические изменения в очаге воспаления
Ишемия, и, в значительно большей степени, венозная гиперемия и стаз, вызывают усиление гликолиза, в результате чего в тканях очага воспаления накапливается молочная кислота; а нарушения липидного обмена ведут к увеличению концентрации свободных жирных кислот и кислых по своей реакции кетоновых тел. Это приводит к тому, что в очаге воспаления накапливается большое количество свободных ионов водорода, то есть развивается состояние ацидоза.
В динамике изменения кислотно-основного состояния при воспалении различают три фазы. В самый начальный период воспалительной реакции развивается кратковременный первичный ацидоз, связанный с ишемией, в процессе которой в тканях увеличивается количество кислых продуктов. При наступлении артериальной гиперемии кислотно-основное состояние в тканях воспалительного очага нормализуется, а затем развивается длительный выраженный метаболический ацидоз, который вначале является компенсированным (происходит снижение щелочных резервов тканей, но их рН не меняется). По мере прогрессирования воспалительного процесса развивается уже некомпенсированный ацидоз вследствие нарастания концентрации свободных водородных ионов и истощения тканевых щелочных резервов. Концентрация водородных ионов повышается тем больше, чем сильнее выражено воспаление. Для гнойного воспаления характерен очень низкий рН (5.0 - 4.0).
В тканях воспалительного очага происходит резкое изменение осмотического и онкотического давления. При альтерации клеток высвобождается большое количество внеклеточного калия. В сочетании с увеличением количества водородных ионов это приводит к гиперионии в очаге воспаления, а последняя вызывает повышение осмотического давления. Накопление полипептидов и других высокомолекулярных соединений приводит к возрастанию онкотического давления. В результате возрастает степень гидратации тканей и их тургор, то есть напряжение, которое при воспалении увеличивается в 7 - 10 раз, что в свою очередь усиливает альтерацию тканей.
Все описанные выше процессы (расширение и увеличение проницаемости микрососудов, физико-химические изменения в очаге воспаления) обеспечивают переход жидкой части крови через сосудистую стенку и образование воспалительного отека. При этом само образование отека является фактором, обеспечивающим его увеличение, так как сдавливание отечной жидкостью микрососудов (в первую очередь – венул) усиливает состояние венозной гиперемии и стаза. Так формируется плазматический компонент экссудации. Одновременно с образованием отека создаются и благоприятные условия для активного перехода лейкоцитов крови через сосудистую стенку и их движения к центру воспалительного очага. Иначе говоря, для клеточной инфильтрации. В значительной степени вопросы клеточной инфильтрации тканей в зоне воспаления будут рассмотрены в разделе «Фагоцитоз». Однако при рассмотрении механизмов экссудации необходимо остановиться на таких явлениях как маргинация, адгезия и диапедез лейкоцитов.
3.2.2.3. Маргинация, адгезия и диапедез лейкоцитов
Нейтрофильные лейкоциты способны за 3 – 12 минут пройти через сосудистую стенку и двинуться к очагу воспаления. Массивное проникновение нейтрофилов через сосудистую стенку приходится на первые 2 часа после начала воспалительного процесса, а их максимальное накопление в очаге воспаления происходит через 4 – 6 часов. Движение нейтрофильных лейкоцитов к очагу воспаления начинается внутри кровеносных сосудов. Приближаясь к участку сосуда, расположенному в непосредственной близости от центра воспалительного очага, лейкоциты замедляют свое движение относительно скорости кровотока и начинают процесс проникновения (диапедеза) через сосудистую стенку. И движение лейкоцитов, и их адгезия к сосудистой стенке, и последующее проникновение через стенку сосуда – это сложный, многоэтапный процесс (Рис.3).
Появление на поверхности лейкоцитов и эндотелиоцитов молекул адгезии начинается только после контакта этих клеток с рядом медиаторов воспаления и цитокинов. До этого контакта молекулы адгезии (L-селектины и бета-2-интегрины в лейкоцитах; Р-селектины и Е-селектины в эндотелиоцитах) содержатся во внутриклеточных гранулах и не функционируют. При этом часть медиаторов и цитокинов действуют только на лейкоциты, а часть – на эндотелиоциты. В частности, такие вещества как лейкотриен В4 и факторы комплемента способствуют освобождению молекул адгезии у лейкоцитов, а интерлейкин-1 (ИЛ-1) и эндотоксины бактерий – у эндотелиоцитов. Другие цитокины (например, фактор некроза опухолей – ФНО) стимулируют адгезивность, как лейкоцитов, так и эндотелиоцитов.

Рис. 3. Маргинация, адгезия и миграции (диапедеза) лейкоцитов
(по W.Bocher, H.Denk, Ph.Heitz)
1 – Р-селектин; 2 – Фактор активации тромбоцитов; 3 – Е-селектин; 4 – иммуноглобу-
линовый комплекс; 5 – хемотаксические цитокины
На первых этапах и движение, и адгезия лейкоцитов и эндотелиоцитов обеспечивается в основном за счет селектинов и, частично, интегринов которые соединяются с соответствующими рецепторами на поверхности лейкоцитов и клеток эпителия сосуда. В последующем происходит «слущивание» (шеддинг) селектинов с поверхности лейкоцитов и их место занимают бета-2-интегрины, резко увеличивающие адгезию лейкоцитов к сосудистой стенке. Дополнительно адгезию усиливают и белки группы иммуноглобулинов, преимущественно экспрессируемые эпителием. Молекулы адгезии оказывают содействие лейкоцитам и при их прохождении через сосудистую стенку. В дальнейшем, при движении лейкоцитов к центру очага воспаления, их «привлечение» обеспечивают молекулы (цитокины) хемотаксиса.
Важнейшей составной частью воспалительного процесса является фагоцитоз, в котором принимают участие как фагоциты крови (лейкоциты), так и фагоциты, находящиеся вне сосудистого русла (например, тканевые макрофаги). Мы, рассматривая процессы фагоцитоза в разделе «Экссудация», основывались на следующих обстоятельствах. Во-первых, диапедез лейкоцитов и их выход в центр очага воспаления – это важнейшая компонента экссудации. Во-вторых, активация фагоцитоза и его завершение в основном осуществляется именно тогда, когда воспалительный процесс в целом проходит стадию экссудации. Рассмотрим механизмы и стадии фагоцитоза.
3.2.2.4. Механизмы фагоцитоза
Определение понятия «фагоцитоз» можно представить в следующем виде:
Фагоцитоз - это процесс поглощения и переваривания клеткой различных корпускулярных агентов (частиц) которые являются или становятся инородными для всего организма или для отдельных его частей[1]/.
В этом определении необходимо подчеркнуть следующие два важных момента. Во-первых, при фагоцитозе происходит процесс поглощения и переваривания не только частиц, изначально являющихся чужеродными для организма, но и тех, которые могут стать таковыми при определенных условиях. Например, микроорганизмы, составляющие нормальную микрофлору кишечника, при их парэнтеральном проникновении в ткани становятся объектами фагоцитоза. Во-вторых, какой-то объект может не быть чужеродным для одной части организма и стать чужеродным для другой. Например, эритроциты в кровеносном русле для организма не чужеродны, но если они попадут в ткани при кровоизлиянии, то могут стать объектами фагоцитоза.
Внутриклеточному захвату и перевариванию могут подвергаться не только корпускулярные агенты, но и жидкие. Захват клетками капель жидкости и использование этих жидкостей в процессах внутриклеточного пищеварения носит название пиноцитоза. Кроме того, чужеродные частицы могут поглощаться фагоцитами и за счет эндоцитоза (так называемое – рецептор-опосредованное взаимодействие).
Явление фагоцитоза было открыто в конце декабря 1882 года И.И.Мечниковым, и в дальнейшем, в результате его работ на протяжении четверти века, было доказано, что фагоцитоз - это один из основных защитных механизмов воспалительной реакции, поскольку он направлен на уничтожение ее причинного фактора.
В фагоцитозе могут принимать участие разнообразные элементы ретикуло-эндотелиальной системы. Но поскольку в данном разделе учебника речь идет о воспалении, фагоцитоз будет рассматриваться применительно только к воспалительной реакции, в которой в качестве основной фагоцитирующей клетки выступают нейтрофильные полиморфноядерные лейкоциты крови и тканевые макрофаги. Нейтрофилы появляются в очаге воспаления чрезвычайно быстро и доминируют там в течение первых 24 часов протекания воспалительной реакции. В дальнейшем в очаге воспаления сосредотачиваются и другие фагоцитирующие клетки (например, тканевые макрофаги, а также моноциты, превратившиеся в макрофаги), и другие иммунокомпетентные клетки (лимфоциты, плазматические клетки и т.п.).
Для более четкого понимания механизмов фагоцитоза следует коротко остановиться на некоторых современных представлениях о строении лейкоцитов, поскольку все, что происходит с фагоцитирующей клеткой, в значительной степени связано с особенностями ее строения.
Согласно этим представлениям лейкоциты имеют клеточный скелет, в состав которого входят микротрубочки, актиновые, миозиновые и промежуточные филаменты. Другими словами, лейкоцит обладает своим опорно-двигательным аппаратом, элементы которого через систему клеточных посредников – мессенджеров связаны с рецепторами, расположенными на поверхности мембраны, в связи с чем цитоскелет лейкоцита в значительной степени определяет и особенности реакции последнего на различные воздействующие на него раздражители. При этом необходимо заметить, что клеточный скелет не представляет собой чего-то постоянного как структурно, так и функционально. Его элементы могут перегруппировываться в зависимости от конкретных условий процесса и требований, которые предъявляются лейкоциту этими условиями. Кроме того, элементы цитоскелета могут менять свое физико-химическое состояние: основной принцип функционирования этого структурного комплекса - процесс обратимой деполимеризации входящих в него белков, регулируемый ионами кальция, кальцийсвязывающим белком - кальмодулином, а также внутриклеточным соотношением цАМФ и цГМФ. Этим определяются важнейшие функции лейкоцита: передвижение, захват чужеродных частиц, внутриклеточное переваривание. Дефекты клеточного скелета лейкоцитов делают их неполноценными, не способными эффективно участвовать в защите организма от факторов, вызывающих воспалительную реакцию.
Процесс фагоцитоза включает в себя несколько стадий.
Первая стадия – стадия маргинации, адгезии и диапедеза лейкоцитов нами уже была рассмотрена выше. Поэтому перейдем к рассмотрению второй стадии фагоцитоза.
Вторая стадия - передвижение фагоцита к объекту фагоцитоза. Это передвижение начинается и поддерживается благодаря тому, что в очаге воспаления образуются вещества, к которым лейкоцит обладает положительным хемотаксисом, то есть при наличии этих веществ лейкоцит начинает двигаться в их сторону. Наиболее известным и изученным хемоаттрактантом для лейкоцитов является интерлейкин 8. Помимо него и другие цитокины обладают хемотаксическими свойствами, например, макрофагальный воспалительный протеин 1-альфа и 1-бета, моноцитарный хемотаксический и активирующий фактор и некоторые другие. Особо можно выделить хемотаксический фактор, образующийся в очаге воспаления в результате окислительного метаболизма арахидоновой кислоты – составной части мембраны лейкоцитов. Это один из лейкотриенов – лейкотриен В4.
Как выяснено, вещества, к которым лейкоцит проявляет положительный хемотаксис, воздействуют на рецепторы его оболочки, в результате чего возникает сенсорный эффект - лейкоцит начинает "чувствовать", "ощущать" эти вещества.
Особенно важное значение в контроле за хемотаксическим процессом имеют циклические нуклеотиды. Показано, что цГМФ повышает чувствительность лейкоцитов к хемотаксическому фактору и усиливает их движение. Противоположным действием обладает цАМФ.
Вещества, к которым у лейкоцитов имеется положительный хемотаксис, меняют физико- химическое состояние их протоплазмы, переводя ее из состояния геля в состояние золя и обратно. Таким образом, какая-то часть протоплазмы лейкоцита становится жидкой и в нее постепенно переливается вся клетка.
Перемещение фагоцита в пространстве осуществляется следующим образом.
Установлено, что протоплазма фагоцита состоит из центрального жидкого слоя (золя) и более плотного наружного - кортикального геля. Под влиянием веществ, к которым лейкоцит обладает положительным хемотаксисом, на переднем полюсе лейкоцита кортикальный гель превращается в золь, то есть, становится более жидким. В эту "разжиженную" часть лейкоцита переливается золь его центральной части, в результате чего лейкоцит укорачивается сзади и удлиняется спереди. Этот процесс по аналогии можно сравнить с выдавливанием зубной пасты из тюбика с той лишь разницей, что и сам "тюбик" (оболочка лейкоцита) устремляется вслед за "пастой" (за протоплазмой).
Существует и другой способ движения фагоцита. Микротрубочки цитоскелета в тот период, когда лейкоцит находится в спокойном состоянии, не имеют четкой ориентации, расположены хаотически и выполняют в основном опорную функцию. Когда же лейкоцит начинает двигаться, эти трубочки меняют свое расположение в цитоплазме и ориентируются точно по направлению движения. Разжиженная часть кортикального геля с переднего полюса лейкоцита засасывается в эти трубочки и с силой выбрасывается из них назад. Возникает реактивная тяга: сами трубочки начинают двигаться в противоположном направлении и толкают лейкоцит вперед. Другими словами, лейкоцит передвигается как ракета. И, наконец, исходя из наличия в лейкоците актин-миозиновой системы, можно предположить, что в нем происходят процессы, аналогичные мышечному сокращению, благодаря чему он и передвигается. Скорость движения лейкоцитов может быть довольно большой. Подсчитано, что за сутки лейкоцит может пройти 5 - 6 см, то есть "добраться" с периферии до центра очень большого по своим размерам воспалительного очага. Передвижение лейкоцитов является энергозависимым процессом, то есть идет с потреблением энергии, причем эту энергию лейкоцит получает от гликолитических реакций. Блокада процессов окислительного фосфорилирования соединениями синильной кислоты не останавливает движения фагоцитов, в то время как монойодацетат, угнетая гликолиз, тормозит этот процесс.[2]
Третья стадия – опсонизация объектов фагоцитоза. Без прохождения этой стадии фагоцитоз не возможен. Опсонизация – это процесс взаимодействия опсонинов (иммуноглобулинов IgG1, IgG3, IgM, белков системы комплемента С3b, C4, C5a, С-реактивного белка) с рецепторным аппаратом инфекционной частицей. На фагоцитах есть рецепторы к Fc-фрагментам иммуноглобулинов и к белкам системы комплемента. В результате инфекционные частицы плотно прикрепляются к оболочке фагоцитов, а иммуноглобулины, белки комплемента и С-реактивный белок служат как бы «мостиком», прочно соединяющим фагоцит и фагоцитируемый объект. Кроме того, определенную роль в прикреплении («прилипании») фагоцита к фагоцитируемому объекту играют некоторые цитокины. Следует указать, что низкий уровень опсонинов в плазме крови, как правило, приводит к тяжелым, длительно текущим инфекционным заболеваниям.
Четвертая стадия - погружение объекта в фагоцит, которая может осуществляться двумя путями. Во-первых, фагоцит, подобно амебе, способен выпускать псевдоподии, которые смыкаются над объектом фагоцитоза, и он оказывается внутри фагоцита. Во-вторых, это погружение может происходить путем инвагинации клеточной оболочки фагоцита: в нем образуется все увеличивающаяся впадина, в которую и погружается объект; затем края впадины смыкаются над объектом, и он оказывается внутри фагоцитирующей клетки. Если же объект по своим размерам очень большой, то он окружается несколькими фагоцитами, которые внедряют в него сливающиеся друг с другом цитоплазматические отростки, и таким путем осуществляется совместный фагоцитоз несколькими фагоцитами одного объекта.
В процессе погружения объекта в фагоцит важную роль играют электрические заряды объекта и фагоцита, интенсивность хемотаксиса и величина поверхностного натяжения в месте соприкосновения фагоцита и фагоцитируемого объекта. Чем ниже этот показатель, тем интенсивнее идет погружение. Поэтому опсонины и бактериотропины, снижающие поверхностное натяжение, способствуют интенсификации фагоцитирования микроорганизмов.
Пятая стадия – переваривание. Вначале живой объект, попавший в фагоцит и находящийся в его пищеварительной вакуоли, должен быть убит. Живые объекты фагоцит не переваривает. Основную роль в гибели живых объектов, попавших в фагоцит, играет резкий сдвиг рН протоплазмы фагоцита в кислую сторону. После того, как объект убит, пищеварительная вакуоль, в которой он находится, сливается с одной или несколькими лизосомами фагоцита, и лизосомные ферменты осуществляют процесс пищеварения в этой полости.
Живой объект может быть фагоцитирован и иным путем: в гранулах лейкоцита содержатся бактерицидные вещества (например, активные кислородные радикалы), которые выбрасываются в окружающую среду, и, таким образом, лейкоцит убивает микроорганизм. Затем осуществляется процесс его погружения и переваривания.
Таковы процессы, лежащие в основе так называемого завершенного фагоцитоза. Однако фагоцитоз протекает по-иному, если микроорганизмы - объекты фагоцитоза либо обладают мощной полисахаридной капсулой (например, микобактерии туберкулеза), защищающей их от кислой реакции среды, либо выделяют вещества, которые препятствуют слиянию лизосом с пищеварительной вакуолью, в результате чего процесс внутриклеточного пищеварения не может быть осуществлен. В этом случае имеет место так называемый незавершенный фагоцитоз. Он заканчивается тем, что через некоторое время живые микроорганизмы либо выбрасываются из фагоцита, либо фагоцит гибнет. Аналогичная ситуация может возникать при некоторых генетически обусловленных дефектах фагоцитарной системы, о чем более подробно рассказывается в разделе учебника, посвященном иммунодефицитным состояниям.
3.2.3. Пролиферация
Острое воспаление завершается фазой пролиферации, хотя пролиферативные процессы в той или иной степени выражены с самого начала воспалительного процесса. Однако интенсивная пролиферация начинается только тогда, когда полностью завершается альтерация и экссудация, а патогенный агент уничтожен или выведен за пределы организма. В противном случае (если патогенный агент не уничтожен) острое воспаление может трансформироваться в хроническое.
Фазе активной пролиферации предшествует период, когда в очаге воспаления начинают вступают в действие противовоспалительные медиаторы. К их числу относятся уже известные нам противовоспалительные цитокины, а также ряд других противовоспалительных биологически активных веществ. Основное место среди них занимают:
- гепарин, который связывает биогенные амины, ингибирует комплемент, является мощным антикоагулянтом, инактивирует кининовые системы;
- ингибиторы протеаз – вещества, подавляющие активность лизосомальных гидролаз и, тем самым, резко снижающие повреждение клеток и тканей;
- антифосфолипазы, ингибирующие фосфолипазу А2 и за счет этого уменьшающие синтез арахидоновой кислоты и ее продуцентов – простагландинов и простациклинов. Следует указать, что выработка антифосфолипаз стимулируется глюкокортикоидными гормонами;
- антиоксиданты – металлосодержащие белки, инактивирующие кислородные радикалы и липоперекиси;
- инактиваторы воспалительных медиаторов, например, гистаминаза и кининаза, разрушающие гистамин и кинины.
В целом, противовоспалительные биологически активные вещества обеспечивают затухание процессов альтерации и экссудации и, тем самым, создают максимально благоприятную ситуацию для запуска механизмов активной пролиферации и репарации.
Репаративные процессы могут идти по двум направлениям: по пути регенерации (замещение погибших клеток клетками точно такого же типа) и по пути фиброплазии (замещение клеточного дефекта соединительной тканью). И регенерация, и фиброплазия осуществляются как за счет усиления пролиферации, так и за счет снижения уровня апоптоза.
Большую роль в регуляции пролиферации играют макрофаги и лейкоциты, которые выделяют ряд медиаторов, стимулирующих, например, трансформацию полибластов в фибробласты.
Ограничивают апоптоз и усиливают пролиферативные процессы вещества, получившие собирательное название «факторы роста». Эти вещества продуцируются макрофагами, тромбоцитами, лимфоцитами и фибробластами. Их действие может уравновешиваться такими цитокинами как фактор некроза опухолей и бета-интерферон, которые могут ингибировать рост ряда клеток в очаге пролиферации.
Как известно, в организме человека и животных постоянно присутствуют вещества, которые получили название «кейлоны». Кейлоны сдерживают клеточный митоз и, тем самым, ограничивают интенсивное размножение клеток, что особенно актуально для некоторых онкологических процессов. На стадии активной пролиферации в очаге воспаления клетки начинают вырабатывать вещества противоположного действия – антикейлоны, стимулирующие клеточное деление.
В регуляции процессов репарации принимают участие и многие гормоны. На первое место здесь нужно поставить тропные гормоны гипофиза и гормоны желез внутренней секреции, регулируемые гипофизом. Активное влияние на рост и размножение фибробластов, клеток паренхиматозных органов, остеобласты и мышечную ткань оказывает соматотропин. Кроме того, под влиянием соматотропина в организме активно синтезируются инсулиноподобные факторы роста - соматомедины и инсулин.
Процессы регенерации костной ткани стимулируются половыми гормонами, а заживление ран – гормонами щитовидной железы.
Таким образом, мы видим, что на заключительной стадии острого воспаления действует большое количество регулирующих факторов, которые обеспечивают как своевременное затухание альтерации и экссудации, так и непосредственно создают благоприятные условия для репарационных и регенерационных процессов.
3.3. Изменения обмена веществ в очаге острого воспаления
С самого начала воспалительной реакции в альтерированных тканях происходят характерные изменения обмена веществ, которые в целом могут быть охарактеризованы как "пожар обмена", то есть как резкая интенсификация всех видов метаболизма. Несколько забегая вперед, укажем, что интенсификация обменных процессов в очаге местного острого воспаления носит, в основном, саногенетический характер, в отличие от синдрома гиперметаболизма, развивающимся при генерализованном воспалении. Этот синдром является крайней степенью патологии и чрезвычайно опасен.
Однако обменные процессы в очаге воспаления меняются не только количественно, но и качественно.
Изменения углеводного, жирового и белкового обмена в очаге воспаления многоплановы и динамичны, поскольку на каждой стадии процесса между метаболическими реакциями возникают новые взаимосвязи, адекватные тем требованиям, которые в каждый конкретный момент предъявляются к клеткам и тканям. Поэтому ниже будут определены только принципиальные изменения этих видов обмена веществ, что необходимо для понимания патогенеза воспалительной реакции.
Углеводный обмен. Начиная с самых ранних стадий воспалительного процесса, в его очаге резко возрастает потребность тканей в кислороде. Несмотря на возникающую артериальную гиперемию, а в дальнейшем - из-за венозной гиперемии, тканям начинает не хватать кислорода. В то же время в воспаленных тканях очень интенсивно используется приносимая в больших количествах с током крови глюкоза. В результате этого усиливается гликолиз, и, как следствие этого, в очаге воспаления происходит накопление больших количеств молочной кислоты.
Характерным для изменений углеводного обмена в очаге воспаления является отсутствие эффекта Пастера, заключающегося в том, что в присутствии кислорода тормозится анаэробное расщепление углеводов. Это обусловлено тем, что при анаэробном расщеплении углеводов на каждую молекулу глюкозы образуется 2 молекулы АТФ, а при аэробном – 38, то есть аэробный путь является гораздо более выгодным для клеток в энергетическом отношении. В условиях развития воспалительной реакции этот механизм нарушается и происходит интенсификация процессов анаэробного расщепления углеводов.
Вам также может быть полезна лекция "Понятие абилитации".
Жировой обмен. В крови, оттекающей от очага воспаления, повышается содержание свободных жирных кислот, так как в воспаленной ткани усиливаются процессы липолиза. Одновременно в этом регионе нарастает количество кетоновых тел, что свидетельствует не только об усилении, но и об извращении жирового обмена.
Белковый обмен. В воспаленных тканях происходит значительное усиление протеолитических процессов, в связи с чем здесь накапливается большое количество аминокислот и полипептидов. Последние в ряде случаев обладают высокой биологической активностью, инициируя ряд метаболических превращений, как в тканях, так и в экссудате.
3.4. Схема патогенеза острого воспаления
Схема патогенеза острого воспаления представлена на Рис. 4. Схема демонстрирует в обобщенно-модульном виде этапность протекания острого воспаления от момента воздействия на ткань повреждающего фактора и до завершения процессов пролиферации, репарации и регенерации. В целом, схема не требует дополнительных комментариев, так как все основные регуляционные механизмы острого воспаления подробно изложены в предшествующем материале. Необходимо, однако, напомнить, что стадии острого воспаления не просто сменяют одна другую, но накладываются друг на друга и могут достигать максимума развития почти одновременно (как это, например, происходит со стадиями вторичной альтерации и экссудации). В свою очередь стадия пролиферации начинает формироваться задолго до окончания стадии экссудации. На схеме активирующие влияния указаны сплошными стрелками, а тормозящие – стрелками пунктирными.
[1]/ И.А.Алов, И.Брауде, М.Е.Аспиз. Основы функциональной морфологии клетки. М. 1966, С.317.
[2] Вполне возможно, что именно отсутствие «эффекта Пастера» в очаге воспаления связано с тем, что лейкоциты для своего передвижения нуждаются в энергии, образующейся именно в процессе анаэробного расщепления углеводов.


















